
Как изменится возбудимость ткани во время возбуждения
В возбудимой ткани невозможно вызвать новый потенциал действия, пока мембрана возбужденного волокна остается деполяризованной в связи с развитием предыдущего потенциала действия. Это связано с тем, что вскоре после возникновения потенциала действия натриевые каналы (или кальциевые каналы, или оба типа каналов) инактивируются, и возбуждающий сигнал любой силы, действующий на ткань в этот момент, не может открыть инактивационные ворота. Единственным условием для их повторного открытия является возврат мембранного потенциала к исходному или близкому к исходному уровню. Затем в течение небольшой доли секунды инактивационные ворота каналов открываются, и становится возможным развитие нового потенциала действия.
Период, в течение которого нельзя вызвать второй потенциал действия даже сильным стимулом, называют абсолютным рефрактерным периодом. Для крупных миелинизированных нервных волокон этот период составляет примерно 1/2500 сек. Легко рассчитать, что такое волокно может максимально передавать около 2500 имп/сек.
Кроме факторов, повышающих возбудимость нервного волокна, существуют так называемые мембраностабилизирующие факторы, способные снижать возбудимость. Например, высокая концентрация ионов кальция во внеклеточной жидкости уменьшает проницаемость мембраны для ионов натрия, снижая возбудимость. В связи с этим ионы кальция называют стабилизатором.
Местные анестетики. К наиболее важным стабилизаторам относят многие вещества, используемые в клинике в качестве местных анестетиков, в состав которых входят прокаин и тетракаин. Большинство из них действуют непосредственно на активационные ворота натриевых каналов, затрудняя их открытие, что сопровождается снижением возбудимости мембран. После снижения возбудимости до уровня, при котором отношение амплитуды потенциала действия к порогу возбуждения (называемое фактором надежности) опускается ниже 1,0, импульсы по анестезированному нерву не проходят.
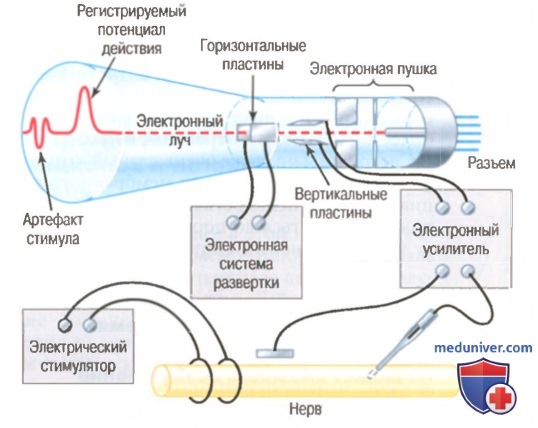
Катодно-лучевой осциллоскоп для регистрации быстрых потенциалов действия.
Катодный осциллограф. Ранее в этой главе мы отмечали, что изменения мембранного потенциала во время генерации потенциала действия происходят чрезвычайно быстро. Действительно, развитие большей части комплекса потенциала действия в крупных нервных волокнах занимает менее 1/1000 сек. На некоторых рисунках данной главы показан электроизмерительный прибор, регистрирующий эти изменения потенциалов. Однако ясно, что реакции любого прибора, способного регистрировать потенциалы действия, должны быть чрезвычайно быстрыми. Для практических целей единственным широко используемым прибором, способным точно реагировать на быстрые изменения мембранного потенциала, является катодно-лучевой осциллоскоп.
На рисунке показаны основные компоненты катодно-лучевого осциллоскопа. Катодно-лучевая трубка состоит из электронной пушки и флюоресцентного экрана, который «бомбардируется» электронами. При ударе электронов о поверхность экрана флюоресцентный материал светится. Если луч электронов перемещается по экрану, вместе с ним движется яркое пятно света, оставляя на экране флюоресцирующую линию.
Кроме электронной пушки и флюоресцентной поверхности, катодно-лучевая трубка снабжена двумя парами электрически заряженных пластин. Одна пара расположена с обеих сторон от электронного луча, а другая — сверху и снизу. Соответствующие электронные усилители изменяют напряжение на этих пластинах таким образом, что электронный луч отклоняется вверх или вниз в ответ на электрические сигналы, приходящие от регистрирующих электродов. Под влиянием внутреннего электронного блока осциллоскопа луч электронов перемещается по экрану горизонтально с постоянной скоростью. При этом регистрируется кривая, которую видно на экране катодно-лучевой трубки рисунка, с временной разверткой по горизонтали и изменениями потенциалов, регистрируемых отводящими электродами, по вертикали.
На левом конце кривой виден небольшой артефакт стимула, связанный с электрическим стимулом, который используется для вызова потенциала действия, справа на кривой — сам потенциал действия.
Видео физиология возбуждения тканей (потенциал покоя, потенциал действия) — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Фазовые изменения возбудимости при потенциале действия
В процессе развития потенциала действия происходит изменение возбудимости возбудимых тканей. Эти изменения носят фазовый характер, связаны с изменениями заряда мембраны и изменениями ее проводимости для ионов (рис. 15).

В период медленной деполяризации (1) начинают открываться натриевые каналы и возбудимость возрастает, что приводит к снижению порога возбуждения. ( Порог возбуждения характеризуется величиной деполяризации от Ео до Ек ). Для открытия дополнительного количества натриевых каналов необходима меньшая сила внешнего раздражителя. Эта фаза повышенной возбудимости (I).
В период быстрой деполяризации (2) все натриевые каналы от- крыты и активированы, входящий ток натрия максимален. Любое внешнее раздражение не создает дополнительной деполяризации. Эта фаза носит название фазы абсолютной рефрактерности (невозбудимости) (II).
В период реполяризации (3) часть натриевых каналов инактивирована медленной h частицей, но часть после инактивации уже перешла в закрытое активированное состояние. Т.е. внешнее воздействие способно перевести их в открытое активированное состояние, а значит вызвать дополнительную деполяризацию мембраны. Сила внешнего раздражения должна быть значительно больше пороговой. Эта фаза носит название относительной рефрактерности (III).
В период следовой деполяризации или следового отрицательного потенциала (4) в связи с остаточно увеличенной натриевой проводимостью мембраны уменьшается порог возбуждения, а возбудимость воз- растает. Эта фаза носит название фазы супернормальной возбудимости или фазы экзальтации (IV). В эту фазу возбуждение может быть вы- звано действием подпорогового стимула.
В период следовой гиперполяризации или следового положительного потенциала (5), обусловленной повышенным током калия, порог возбуждения увеличивается, а возбудимость уменьшается. Эта фаза субнормальной возбудимости (V). Возбуждение может быть вызвано только сверхпороговыми раздражителями.
Изменения возбудимости при возбуждении
Используя тестирующие стимулы в различные фазы развития потенциала действия, можно проследить временной ход изменений возбудимости, сопровождающих возбуждение. На рис. 21 видно, что во время развития локального ответа возбудимость повышается (мембранный потенциал приближается к критическому уровню деполяризации); во время пика потенциала действия мембрана утрачивает возбудимость, которая постепенно восстанавливается после окончания пика. Период полной невозбудимости получил название «фаза абсолютной рефрактерности» . Она обусловлена практически полной инактивацией натриевых каналов и повышением калиевой проводимости. Реполяризация мембраны ведет к реактивации натриевых каналов и снижению калиевой проводимости.

Это период так называемой относительной рефрактерности. В данную фазу возбудимость постепенно возрастает.
В нервных волокнах длительность фазы относительной рефрактерности составляет 5—10 мс . При наличии следовой деполяризации фаза относительной рефрактерности сменяется фазой повышенной возбудимости («супернормальности»). В этот период пороговый потенциал ∆V и соответственно порог раздражения снижены по сравнению с исходными значениями, поскольку мембранный потенциал ближе к критической величине, чем в состоянии покоя. В быстрых двигательных волокнах теплокровных животных фаза следового повышения возбудимости продолжается до 30 мс (рис. 22).
Следовая гиперполяризация, напротив, сопровождается снижением возбудимости. По своему механизму это снижение возбудимости сходно с тем, которое имеет место при анэлектротоне: ∆V увеличен за счет удаления мембранного потенциала Е от критической величины Ек. Сходство это, однако, неполное: при анэлектротоне gK низко, а во время следовой гиперполяризацип g к увеличено, что также повышает порог раздражения.
Читайте также: Характеристика механической ткани кратко
1)Понятия раздражения, раздражимости, возбуждения, возбудимости. Виды раздражителей: адекватные, неадекватные.
Раздражимость-способность живых клеток реагировать изменением обмена веществ в ответ на действие раздражителей.
Раздражние-реакция живой клетки,ткани,органа на внешнее воздействие.
Возбудимость- свойство клеток отвечать на раздражение возбуждением.
Возбуждение-ответ ткани на ее раздражение, проявляющийся в специфической для нее функции(проведение возбуждения, сокращение мышцы, секреция железы) и неспецифических реакциях(генерация потенциала действия, метаболические изменения).
Адекватными называются раздражители, на действие которых определенный вид организмов, орган или живая ткань приспособились соответственно реагировать в естественных условиях на протяжении многих тысячелетий исторического развития. Например, для глаза адекватны световые раздражители, для уха — звуковые и т. д.
Неадекватными называются раздражители, не соответствующие строению и функции воспринимающего органа, например, механические и электрические раздражители для глаза. Они также вызывают изменение в нем обмена веществ, но отличающееся от вызванного действием адекватного раздражителя, и поэтому не могут вызвать точно такие же изменения функции, как адекватный раздражитель.
2)Изменение возбудимости при возбуждении. Прямое и непрямое раздражение мышц.
Изменение возбудимости при возбуждении. При возбуждении возбудимость изменяется пофазно. 1) фаза первичной экзальтации — возбудимость выше нормы, реакция на порошковый и подпороговый раздражитель (соответствует 1 фазе ПД — медленной деполяризации) 2) фаза абсолютной рефрактерности — ответная реакция на раздражитель отсутствует, что обусловлено инактивацией натриевых каналов (соответствует быстрой деполяризации ПД) 3) фаза относительной рефрактерности — возбудимость восстанавливается, и ответная реакция становится возможной только при действии раздражителя надпороговой силы, что обусловлено выходящим калиевым током (соответствует фазе реполяризации) 4) фаза вторичной экзальтации — ответная реакция на подпороговый раздражитель (соответствует следовой деполяризации) 5) фаза субнормальной возбудимости — возбудимость ниже нормы, ответ возможен на действие надпороговой силы (соответствует следовой гиперполяризации)
Прямым раздражением называется непосредственное действие раздражителя на орган, например, раздражение электрическим током мышцы, выпрепарованной из организма. Непрямое раздражение производится действием раздражителя на рецепторы—специальные органы, расположенные на внешней поверхности организма или внутри его и воспринимающие раздражение, например, глаза, уши, органы обоняния, вкуса, рецепторы кожи, мышц, суставов, сухожилий, внутренних органов. При непрямом раздражении органа волны, или импульсы возбуждения из рецептора сначала поступают по центростремительным нервам в центральную нервную систему, а затем уже направляются в орган по центробежным нервам и вызывают его функцию или вызывают образование в некоторых органах веществ, действующих через кровь.
3)Критерии оценки возбудимости: пороговая сила, пороговое время, критический уровень деполяризации.
Пороговая сила – это наименьшая сила раздражителя, способная вызвать ПД при неограниченном во времени действии раздражителя. При использовании в качестве раздражителя электрический ток, его пороговая сила равна 1 реобазе. Если возбудимость ткани высока, пороговая сила раздражителя мала.
Пороговое время – это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение. Хронаксия – наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение
Критический уровень деполяризации — пороговый уровень деполяризации плазмалеммы возбудимой клетки. Уровень деполяризации, при достижении которого возникает потенциал действия. В пределах от потенциала покоя до критического уровня деполяризации между интенсивностью раздражителя и уровнем деполяризации существует приблизительно линейная зависимость. При деполяризации достигающей критического уровня такая зависимость отсутствует (закон «все или ничего»).
4.5. Изменения возбудимости клетки во время ее возбуждения. Лабильность
А. Возбудимость клетки во время ее возбуждения быстро и сильно изменяется. Различают несколько фаз изменения возбудимости, каждая из которых строго соответствует определенной фазе ПД и так же, как и фазы ПД, определяется состоянием проницаемости клеточной мембраны для ионов. Схематично эти фазы представлены на рис. 4.6.
1. Кратковременное повышение возбудимости в начале развития ПД, когда уже возникла некоторая деполяризация клеточной мембраны. Если деполяризация не достигает критической величины, то регистрируется локальный потенциал. Если же деполяризация достигает Екр, то развивается ПД. Возбудимость повышена потому, что клетка час-
тично деполяризована, мембранный потенциал приближается к критическому уровню и, когда деполяризация достигает примерно 50 % пороговой величины, начинают открываться потенциалчувствительные быстрые Na-каналы. При этом достаточно небольшого увеличения силы раздражителя, чтобы деполяризация достигла Екр, при которой возникает ПД.
Абсолютная рефрактерная фаза — это полная невозбудимость клетки, (возбудимость равна нулю), она соответствует пику ПД и продолжается 1—2 мс; если ПД более про должителен, то более продолжительна и аб солютная рефрактерная фаза. Клетка в этот период времени на раздражения любой силы не отвечает. Невозбудимость клетки в фазах деполяризации и восходящей части инверсии объясняется тем, что потенциалзависимые т-ворота Na-каналов уже открыты и Na + бы стро поступает в клетку по всем открытым каналам. Те ворота Na-каналов, которые еще не успели открыться, открываются под влия нием деполяризации — уменьшения мем бранного потенциала. Поэтому дополнитель ное раздражение клетки относительно дви жения Na + в клетку ничего изменить не может. Именно поэтому ПД либо совсем не возникает при раздражении, если оно мало, либо является максимальным, если действует раздражение достаточной силы (пороговой или сверхпороговой). В период нисходящей части фазы инверсии клетка невозбудима по тому, что закрываются инактивационные h-ворота Na-каналов, в результате чего кле точная мембрана непроницаема для Na + даже при сильном раздражении. Кроме того, в этот период открываются (уже в большом ко личестве) К-каналы, К + быстро выходит из клетки, обеспечивая нисходящую часть фазы инверсии и реполяризацию. Абсолютная рефрактерная фаза продолжается и в период реполяризации клетки до достижения уровня мембранного потенциала Екр + 10 мВ. Абсо лютный рефрактерный период ограничивает максимальную частоту генерации ПД. Если абсолютный рефрактерный период заверша ется через 2 мс после начала ПД, клетка может возбуждаться с частотой максимум 500 имп/с. Существуют клетки с еще более коротким рефрактерным периодом, в кото рых возбуждение может в крайних случаях повторяться с частотой 1000 имп/с. С такой частотой могут возбуждаться нейроны рети кулярной формации ЦНС, толстые миелино- вые нервные волокна.
Относительная рефрактерная фаза — это период восстановления возбудимости
клетки, когда сильное раздражение может вызвать новое возбуждение (см. рис. 4.6, Б-3). Относительная рефрактерная фаза соответствует конечной части фазы реполяризации (начиная от Екр. ± 10 мВ) и следовой гиперполяризации клеточной мембраны, если она имеется. Пониженная возбудимость является следствием все еще повышенной проницаемости для К + и избыточного выхода его из клетки. Поэтому, чтобы вызвать возбуждение в этот период, необходимо приложить более сильное раздражение, так как выход К + из клетки препятствует ее деполяризации. Кроме того, в период следовой гиперполяризации мембранный потенциал больше и, естественно, дальше отстоит от критического уровня деполяризации. Если реполяризация в конце пика ПД замедляется (см. рис. 4.4, А), то относительная рефрактерная фаза включает и период замедления реполяризации, и период гиперполяризации, т.е. продолжается до возвращения мембранного потенциала к исходному уровню после гиперполяризации. Продолжительность относительной рефрактерной фазы вариабельна, у нервных волокон она невелика и составляет несколько мс.
Читайте также: Кресло для руководителя everprof ultra t черное сетка ткань металл
4. Фаза экзальтации — это период повышенной возбудимости. Он соответствует следовой деполяризации. В некоторых клетках, например в нейронах ЦНС, возможна частичная деполяризация клеточной мембраны вслед за гиперполяризацией. Очередной ПД можно вызывать более слабым раздражением, поскольку мембранный потенциал несколько ниже обычного и оказывается ближе к критическому уровню деполяризации, что объясняют повышенной проницаемостью клеточной мембраны для ионов Na + . Скорость протекания фазовых изменений возбудимости клетки определяет ее лабильность.
Б. Лабильность, или функциональная подвижность (Н.Е.Введенский), — это скорость протекания одного цикла возбуждения, т.е. ПД. Как видно из определения, лабильность ткани зависит от длительности ПД. Это означает, что лабильность, как и ПД, определяется скоростью перемещения ионов в клетку и из клетки, которая в свою очередь зависит от скорости изменения проницаемости клеточной мембраны. При этом особое значение имеет длительность рефрактерной фазы: чем больше рефрактерная фаза, тем ниже лабильность ткани.
Мерой лабильности является максимальное число ПД, которое ткань может воспроизвести в 1 с. В эксперименте лабильность исследуют с помощью регистрации максимального
числа ПД, которое может воспроизвести клетка при увеличении частоты ритмического раздражения.
Лабильность различных тканей существенно различается. Так, лабильность нерва равна 500—1000, мышцы — около 200, нервно-мышечного синапса — порядка 100 импульсов в секунду. Лабильность ткани понижается при длительном бездействии органа и при утомлении, а также в случае нарушения иннервации.
Следует отметить, что при постепенном увеличении частоты ритмического раздражения лабильность ткани повышается, т.е. ткань отвечает более высокой частотой возбуждения по сравнению с исходной частотой. Это явление открыто А.А.Ухтомским и называется усвоением ритма раздражения.
4.6. ОЦЕНКА ВОЗБУДИМОСТИ КЛЕТКИ. АККОМОДАЦИЯ. ИСПОЛЬЗОВАНИЕ ЭЛЕКТРИЧЕСКОГО ТОКА В МЕДИЦИНЕ
Возбудимость клетки изменяется не только в процессе возбуждения, но и при изменении химического состава внеклеточной жидкости, например, в результате длительной высокой активности клеток, отклонения показателей внутренней среды в патологических случаях. При снижении концентрации ионов Na + вне клетки этот ион в меньшем количестве входит в клетку, в результате чего снижается ее возбудимость из-за гиперполяриза-ции клетки. Это наблюдается, например, при бессолевой диете, при этом может развиваться мышечная слабость. Повышение внеклеточной концентрации Na + вызывает противоположный эффект, например усиление тонуса сосудов вследствие возрастания возбудимости нервно-мышечных элементов. Возбудимость различных тканей сама по себе различна — у нервных клеток выше, чем у мышечных, что используется в клинической практике, например, при выяснении причины двигательных нарушений. Показателями состояния возбудимости ткани являются пороговый потенциал, пороговая сила, пороговое время.
А. Пороговый потенциал (ДУ) — это минимальная величина, на которую надо уменьшить мембранный потенциал покоя, чтобы вызвать возбуждение (ПД). AV и возбудимость клеток находятся в обратных соотношениях: небольшая величина AV свидетельствует о высокой возбудимости клетки. Если, например, уменьшение мембранного потенциала (частичная деполяризация) на 5—
10 мВ вызывает возникновение ПД, то возбудимость клетки высока. Напротив, большой &V (30—40 мВ) свидетельствует о более низкой возбудимости клетки. Однако во всех случаях ПД возникает только при достижении критического уровня деполяризации клеточной мембраны (Екр.).
Критический уровень деполяризации Екр_ (КУД) — это минимальный уровень деполяризации клеточной мембраны, при которой возникает ПД. Дальнейшее раздражение клетки и искусственное снижение ПП ничего не изменяют в процессе возникновения ПД, поскольку деполяризация клетки, достигнув критического уровня, сама по себе ведет к открытию потенциалзависимых т-ворот Na-каналов, в результате чего Na + устремляется в клетку, ускоряя деполяризацию независимо от действия раздражителя. Критический уровень деполяризации клеточной мембраны обычно составляет около —50 мВ. При величине ПП, например, —60 мВ (Ео) деполяризация — уменьшение ПП на 10 мВ приведет к достижению Екр. (—50 мВ) и возникнет ПД. Если ПП равен -90 мВ, то для вызова ПД надо снизить ПП на 40 мВ. В последнем случае возбудимость клетки значительно ниже.
Соотношения между AV, Ео и Е^, показаны на рис. 4.7 — наибольшая возбудимость npvi надме-йкшетА &V, т\\мел\ъшл% ъо^мл-мость при наибольшем А\. AV мало зависит от критического уровня деполяризации (Екр), но существенно — от ПП клетки (Ео), поскольку Екр, как отмечалось выше, — величина довольно постоянная.
Величина ПП изменяется в различных условиях деятельности клетки, вследствие этого колеблется и ее возбудимость, например при изменении концентрации Са 2+ , рН среды. Когда концентрация Са 2+ в среде повышается, клетка становится менее возбудимой, поскольку возрастает мембранный потенциал, вследствие чего Ео удаляется от Екр,, а когда концентрация Са 2+ снижается, возбудимость клетки возрастает, так как мембранный потенциал уменьшается, Ео приближается к Екр. Такое повышение возбудимости лежит в основе синдрома тетании, связанного с дефицитом Са^ + в крови (см. раздел 4.3.1). Изменения содержания ионов Н + в среде действуют на возбудимость нейронов так же, как изменения концентрации Са 2+ , что в обоих случаях объясняется изменением
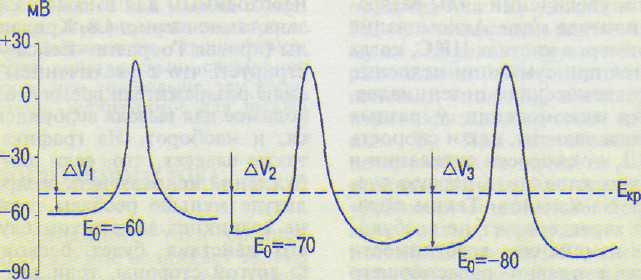
Рис. 4.7. Зависимость возбудимости клетки от величины мембранного потенциала (Ео) при одинаковой величине критического уровня деполяризации мембраны (Екр).
величины Ео. Однако если мембранный потенциал снижается медленно ниже Екр. (—50 мВ), например, в условиях гипоксии, при действии миорелаксантов типа сукцинилхо-лина, то клетка становится невозбудимой вследствие инактивации № + -каналов и невозможности достичь Екр.
Несмотря на то что AV является наиболее точным показателям состояния возбудимости клетки, используется он в эксперименте из-за сложности процедуры реже, чем другие показатели. Чаще всего возбудимость оценивается по пороговой силе раздражителя.
Б. Пороговая сила — это наименьшая сила раздражителя, способная вызвать возбуждение (ПД) при неограничении ее действия во времени (рис. 4.8). Сила раздражителя — понятие собирательное, оно отражает степень выраженности раздражающего воздействия стимула на ткань. Например, сила электрического тока выражается в амперах (А), температура среды — в градусах Цельсия (°С), концентрация химического вещества — в миллимолях на 1 л (ммоль/л), сила звука — в децибелах (дБ) и т.д. При использовании в качестве раздражителя электрического тока предложенное определение пороговой силы совпадает с понятием «реобаза». Реобаза — наименьшая сила тока, способная вызвать импульсное возбуждение. Если возбудимость ткани высока, пороговая сила раздражителя мала. Чем выше возбудимость, тем ниже пороговая сила. Большая пороговая сила свидетельствует о низкой возбудимости ткани. При внутриклеточном раздражении пороговая сила электрического тока для различных клеток равна 1(Г 7 —10
Читайте также: Лента клеющаяся для ткани утюгом
9 А. При медленно нарастающей силе раздражителя возбуждение может не возникнуть даже при достижении большой
его силы, значительно превосходящей пороговую. Это свидетельствует о том, что возбудимость ткани в таких условиях уменьшается — возникает явление аккомодации.

Аккомодация — это понижение возбудимости ткани и амплитуды ПД вплоть до полного его отсутствия при медленно нарастающем стимуле (малая крутизна). Главной причиной аккомодации является инактивация Na-каналов, возникающая при медленной деполяризации клеточной мембраны — в течение 1 с и более. Клетка теряет возбудимость, если закрывается около 50 % инакти-вационных h-ворот (в покое h-ворота в основном открыты, закрыто около 20 %). Меньшую роль играет активация К-каналов. Возникающая частичная деполяризация клетки при медленно нарастающей силе стимула ведет к уменьшению мембранного потенциала. Поэтому если и возникает ПД при
О 0,4 0,8 Рис. 4.8. Кривая «сила
На ординате — сила, на абсциссе — длительность раздражения. Точка А отражает пороговую силу (реобазу) и пороговое (полезное) время, точка Б — удвоенную реобазу и хронаксию.
дальнейшем резком увеличении силы раздражителя, то его амплитуда мала. Аккомодация развивается, например, в клетках ЦНС, когда они деполяризуются при суммации медленно нарастающих синаптических потенциалов. Скорость развития аккомодации у разных тканей различна, она зависит, как и скорость возникновения ПД, от скорости активации и инактивации ионных каналов, в первую очередь инактивации Na-каналов. Таким образом, аккомодация характеризует не возбудимость ткани, а изменение возбудимости ткани при действии плавно нарастающего раздражителя. Поэтому при определении возбудимости ткани в качестве раздражителя необходимо использовать прямоугольные импульсы. В этом случае скорость нарастания стимула и активация Na-каналов опережают скорость аккомодации (инактивации Na-каналов), что и приводит к возникновению ПД. Важным условием, обеспечивающим возникновение возбуждения при действии раздражителя, является его длительность. Поэтому для оценки свойств возбудимой ткани вводится еще одно понятие — пороговое время.
В. Пороговое время — это минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение (см. рис. 4.8 — проекция точки А на абсциссу). Пороговое время называют также полезным временем, так как раздражитель обеспечивает деполяризацию только до критического уровня (Екр). Далее ПД развивается независимо от действия раздражителя, дальнейшее раздражение уже становится ненужным — бесполезным. В эксперименте в клинической практике для оценки свойств возбудимой ткани чаще используют не пороговое время, а хронаксию. Это связано с тем, что определение порогового времени затруднено (пологая часть кривой, см. рис. 4.8). Хронаксия — наименьшее время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение (см. рис. 4.8 — проекция точки Б на абсциссе). Хронаксия соответствует более крутой части кривой сила — время, она колеблется от сотых долей до сотен миллисекунд. Измерение хронаксии в клинической практике позволяет уточнить характер повреждений мышцы при травмах. В норме определяется фактически хронаксия нервных волокон, так как возбудимость их выше. В случае повреждения нерва и его перерождения определяют истинную хронаксию мышцы, которая намного превышает таковую до травмы.
Взаимозависимость между сверхпороговой силой раздражителя и временем его действия,
необходимым для вызова возбуждения, показана также на рис. 4.8. Кривая в виде гиперболы (кривая Гоорвега—Вейса—Лапика) демонстрирует, что с увеличением сверхпороговой силы раздражителя время его действия, необходимое для вызова возбуждения, уменьшается, и наоборот. Из графика (правая часть) также следует, что, если для получения возбуждения использовать раздражитель по амплитуде меньше реобазы, возбуждение ткани не возникнет даже в том случае, если время его действия будет бесконечно большим. С другой стороны, если для получения возбуждения использовать раздражитель, длительность которого будет меньше некоторого критического интервала (левая часть графика), возбуждение ткани также не будет получено даже в том случае, если сила раздражителя будет бесконечно большой. Поэтому высокочастотный переменный ток (>10 кГц) опасности для организма не несет: при сверхкоротком воздействии на ткань импульс электрического тока дает лишь тепловой эффект, что используется в клинической практике для глубокого прогревания тканей при различных патологических процессах. Электрический ток с частотой от 0,5 до 1 мГц также может использоваться в лечебных целях, в основном для прогревания тканей. Низкочастотный переменный синусоидальный ток (50 Гц) стимулирует возбудимые ткани. Стимулы синусоидального тока частотой 50 Гц большого напряжения опасны для жизни: они могут вызвать фибрилляцию сердца с летальным исходом (относительный рефрактерный период миокарда представляет собой в этом случае фазу повышенной уязвимости).
В учебниках и руководствах по физиологии нередко встречаются термины «порог раздражения» и «порог возбуждения», под которым понимают минимальную интенсивность или минимальную энергию раздражения, способную вызвать возбуждение. Следует заметить, что понятие «интенсивность раздражения» не определено в количественном отношении. Понятие «порог раздражения» или «порог возбуждения» как минимальная энергия без указания времени действия раздражителя тоже не позволяет определить возбудимость ткани, поскольку при длительном действии слабого (подпороговой силы) раздражителя можно израсходовать большое количество энергии, но возбуждения ткани так и не вызвать. В этом случае деполяризация не достигнет критического уровня (Екр.). Кроме того, возникнет явление аккомодации — инактивация Na-каналов, которое, как известно, развивается быстро. Необоснованно также рассматривать термин «пороговая сила» как величину, зависящую от времени ее действия, что нередко допускается. Пороговая сила не может за-
висеть от времени действия — она зависит только от возбудимости ткани. При определении пороговой силы время ее действия не ограничивают. Сверхпороговая сила действительно связана со временем ее действия: чем она больше, тем короче время ее действия, необходимое для вызова возбуждения.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом