
4. Транспортные липопротеиды крови: состав, строение, классификация функции, диагностическое значение определения.
Транспорт липидов в организме идет двумя путями:
1) ЖК транспортируются в крови с помощью альбуминов;
2) липиды транспортируются в крови в составе липопротеинов.
ЛП – это надмолекулярные комплексы сферической формы, состоящие из липидов, белков и углеводов. ЛП имеют гидрофильную оболочку и гидрофобное ядро. В гидрофильную оболочку входят белки и амфифильные липиды. В гидрофобное ядро входят гидрофобные липиды и эфиры. ЛП хорошо растворимы в воде.
В организме синтезируются несколько видов ЛП, они отличаются химическим составом, образуются в разных местах и осуществляют транспорт липидов в различных направлениях:
1) хиломикроны образуются в энтероцитах, транспортируют к тканям экзогенные липиды пищи;
2) ЛПОНП (липопротеины очень низкой плотности) образуются в гепатоцитах, транспортируют к тканям эндогенные липиды печени;
3) ЛПП (липопротеины промежуточной плотности) образуются в крови из ЛПОНП, транспортируют к тканям эндогенные липиды печени;
4) ЛПНП (липопротеины низкой плотности) образуются в крови из ЛППП, транспортируют холестерин в ткани;
5) ЛПВП (липопротеины высокой плотности) образуются в гепатоцитах, удаляет избыток холестерина из тканей, донор апо А, С, Е.
5. Катаболизм триглицеридов в белой жировой ткани: реакции, механизмы регуляции активности липазы жировых клеток, роль гормонов, значение.
Жиры депонируются в специализированных клетках жировой ткани – адипоцитах. Мобилизация депонированных жиров происходит путем их гидролиза до жирных кислот и глицерина липазами жировых клеток. ЖК поступают в кровь, где образуют нековалентные соединения с альбумином, и в такой форме транспортируются по кровеносному руслу. Глирерин транспортируется в растворенном состоянии и улавливается главным образом печенью; в печени глицерин превращается в а-глицоролфосфат, который может вступать в реакции глюконеогенеза или окисляться в реакциях гликолиза и общего пути катаболизма. Адреналин активирует мобилизацию депонированных жиров, действуя по тому же механизму, как и в случаях мобилизации гликогена: синтез цАМФ, активация протеинкиназы и фосфорилирование липазы. Катаболизм триглицеридов актуален при длительном голодании и физических нагрузках.
6. Биосинтез триглицеридов: реакции, механизмы регуляции, роль гормонов, значение.
Синтез триглицеридов происходит из глицерина и жирных кислот (главным образом стеариновой, пальмитиновой и олеиновой). Путь биосинтеза триглицеридов в тканях протекает через образование α-глице-рофосфата (глицерол-3-фосфата) как промежуточного соединения.
В почках, а также в стенке кишечника, где активность фермента глицеролкиназы высока, глицерин фосфорилируется за счет АТФ с образованием глицерол-3-фосфата: [глицерин+ АТФ (глицеролкиназа)→ глицерол-3-фосфат + АДФ].
В жировой ткани и мышцах вследствие очень низкой активности глицеролкиназы образование глицерол-3-фосфата в основном связано с процессами гликолиза и гликогенолиза: [дигидроксиацетонфосфат + НАДН2 (глицеролфосфатдегидрогеназа)↔ глицерол-3-фосфат + НАД].
Образовавшийся тем или иным путем глицерол-3-фосфат последовательно ацилируется двумя молекулами КоА-производного жирной кислоты, в результате образуется фосфатидная кислота: [глицерол-3-фосфат (глицеролфосфатацилтрансфераза)↔ фосфатидная к-та].
Далее фосфатидная к-та гидролизуется фосфатидатфосфогидролазой до 1,2-диглицерида (1,2-диацилглицерола): [фосфатидная к-та + Н2О (фосфатидатфосфогидролаза)→ 1,2-диглицерид].
Затем 1,2-диглицерид ацилируется третьей молекулой ацил-КоА и превращается в триглицерид (триацилглицерол): [1,2-диглицерид (диацилглицеролацилтрансфераза)↔ Триглицерид]. Синтез триглицеридов (триацилглицеролов) в тканях происходит с учетом двух путей образования глицерол-3-фосфата и возможности синтеза триглицеридов в стенке тонкой кишки из β-моноглицеридов, поступающих из полости кишечника в больших количествах после расщепления пищевых жиров.
Катаболизм липидов

Внешний обмен липидов
Источник липидов в пище тебе тоже хорошо известен. Это все, что вкусно и вредно. Ну и рыба еще. Расщепление липидов происходит в двенадцатиперстной кишке, но тут уже есть чуть больше нюансов, чем с глюкозой. Липиды очень гидрофобны и собираются, как бабули на скамейке, в мицеллы, обращаясь кнаружи гидрофильными участками молекул. И в этот интимный круг никак не пробиться, если предварительно не эмульгировать их. Для этого в организме придуманы желчные кислоты. Они способствуют распаду липидных капель на миллиарды еще более мелких капель, что делает их доступными для действия панкреатических ферментов [1–3].
В панкреатическом соке, если долго вглядываться, можно найти липазу. Она расщепляет боковые жирные кислоты ТАГ, оставляя после себя 2-моноацилглицерол (2-МАГ).
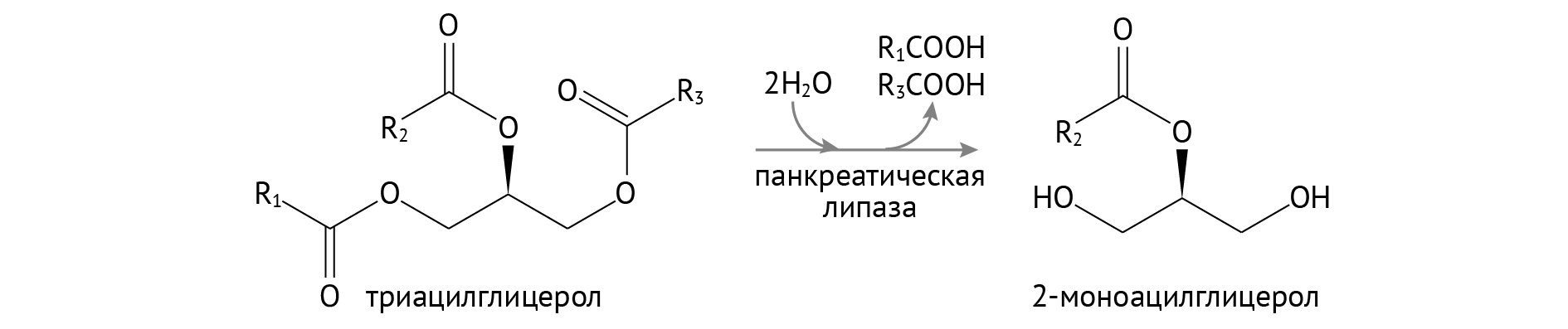
Хотя в ряде случаев все это разрезается до жирных кислот и глицерина.
Читайте также: Бюрократ детское кресло бюрократ ch w797 сетка ткань крестовина пластик
Продукты расщепления просто превосходно всасываются в энтероциты. Но выбросить жирные кислоты в кровь просто так нельзя. В энтероцитах происходит их ресинтез. Во многом (как ни странно) он повторяет процесс расщепления.
Ресинтез. Чтобы присоединить жирную кислоту к глицерину/2-МАГ, необходимо навести на кислоту марафет. Косметическим элементом для нее служит S-KoA-группа.
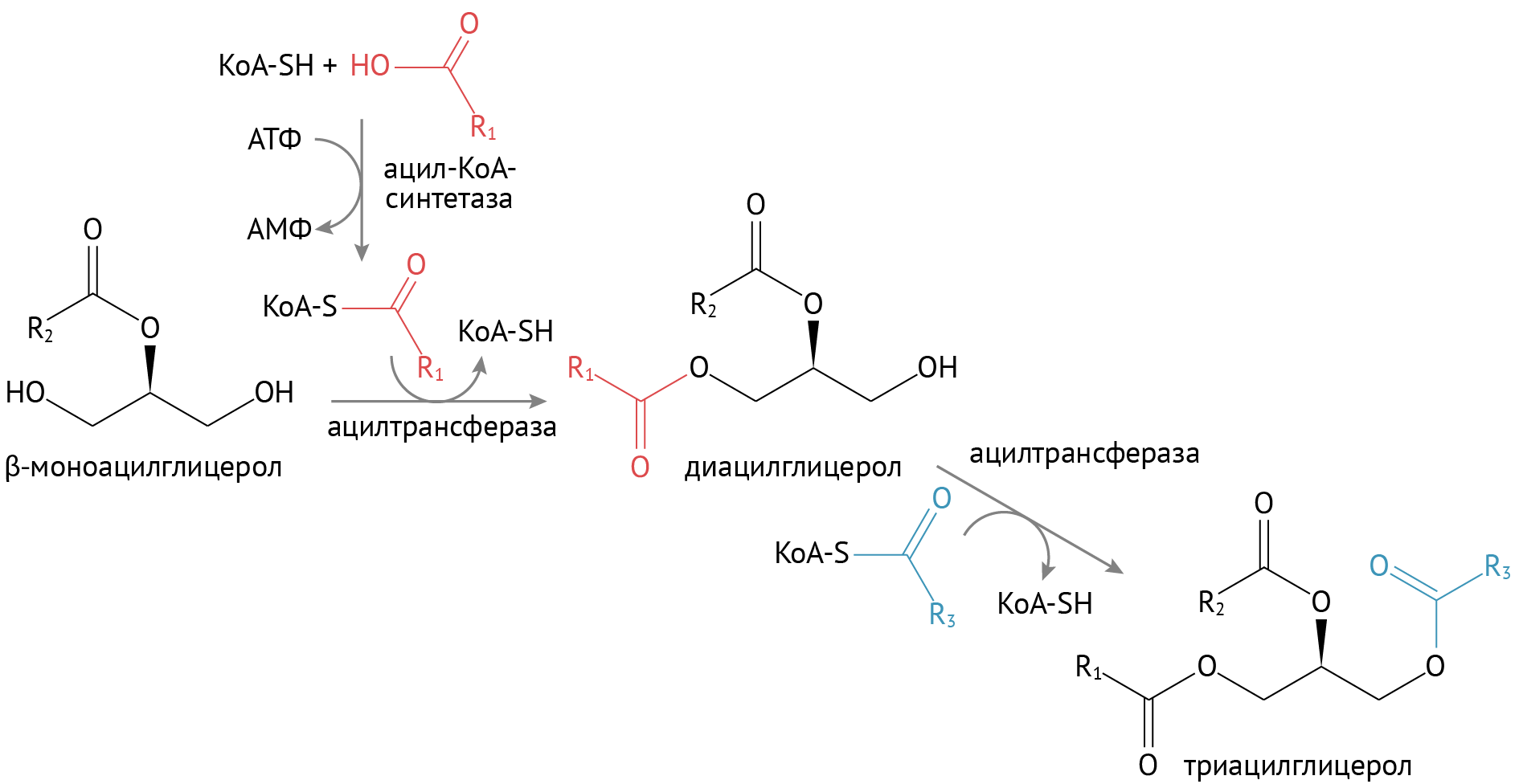
В энтероците собираются еще и фосфолипиды — не менее важные для организма (я бы сказал, самые важные) липиды. Их синтез происходит просто: к диацилглицеролу присоединяется холин.
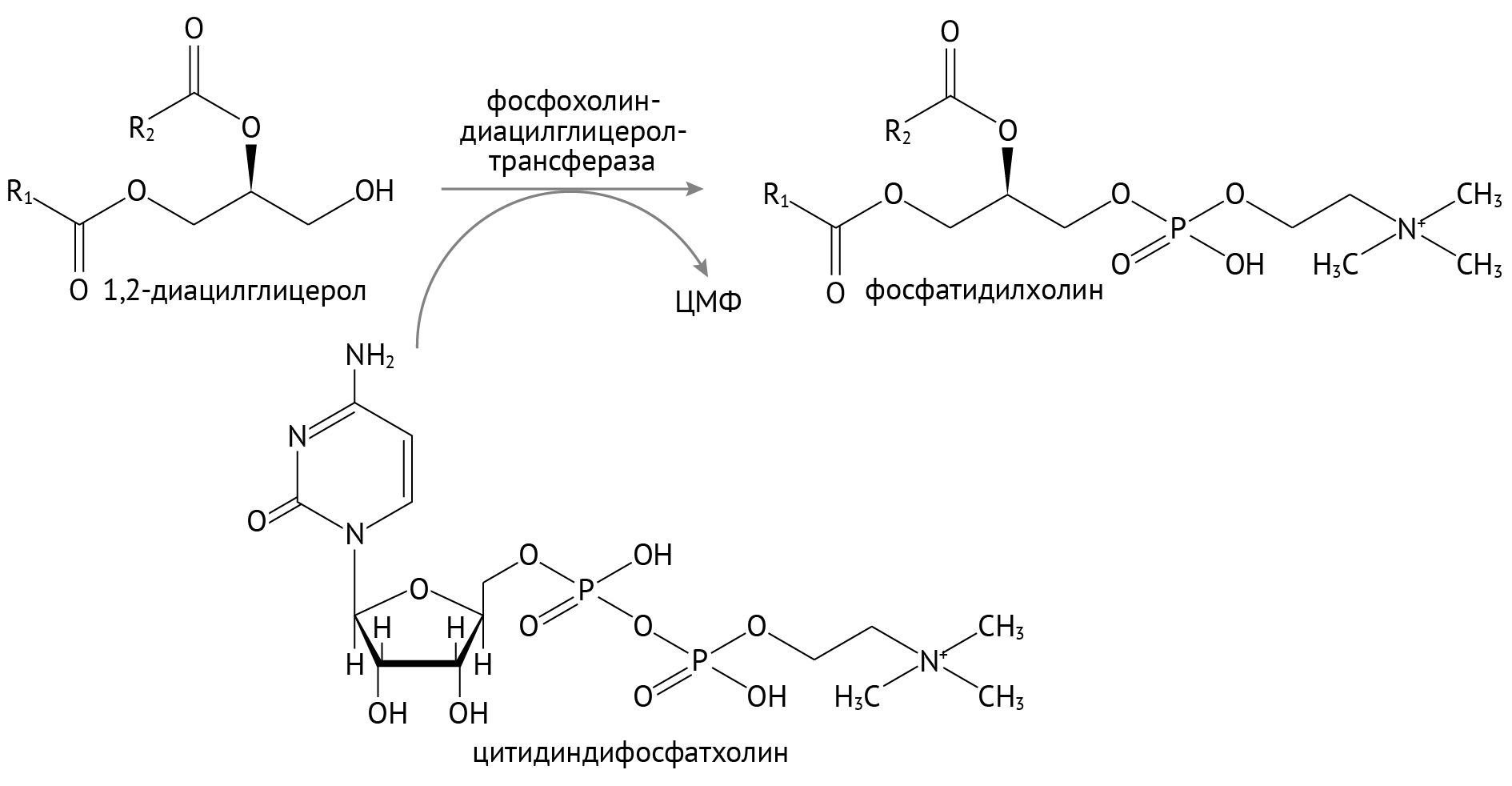
Весь этот комок липидов сбивается в кучу под названием хиломикроны.
Путешествие липидов от кишки до клеток
Липиды, всосавшиеся в кишке, достигают системы верхней полой вены. Но если ты помнишь анатомию, это для тебя не должно быть сюрпризом. Все объясняется просто: хиломикроны всасываются в систему лимфатических капилляров и сосудов. Выбросить их в воротную вену будет насилием над печенью, поэтому мы насилуем лимфатическую дренажную систему. Путь хиломикронов в анатомическом отношении принимает следующий характер. Лимфатические капилляры кишки → лимфатические сосуды брюшной полости → левый грудной лимфатический проток → венозный угол (место слияния левой внутренней яремной и левой подключичной вен, а также место впадения грудного лимфатического протока) → плечеголовная вена → система верхней полой вены [1].
Попадая в систему легочных сосудов, а затем в малый круг, хиломикроны разносятся по тканям. Во многих тканях (особенно в жировой) присутствует фермент липопротеидлипаза. Этот товарищ расщепляет хиломикроны, вычленяя оттуда триглицериды, фосфолипиды, жирные кислоты и вот это вот все. Те благополучно достигают клеток и спокойно попадают внутрь, в цитозоль. Важно отметить, что для активации липопротеинлипазы тканей необходим апопротеин С — II [2].
Хиломикроны приобретают его при взаимодействии с липопротеидами высокой плотности (ЛПВП) в плазме крови.
P. S. Oбмен липопротеидов мы пока опустим и подробно разберем его в другой лекции. А заодно узнаем о биохимических предпосылках атеросклероза — настоящей чумы нашего времени.
Распад триацилглицеридов в тканях
Триацилглицериды (ТАГ) являются запасной формой энергии в силу того, что при окислении жирных кислот получается просто огромнейшее ее количество. Но чтобы окислить, их нужно достать из лап глицерина.
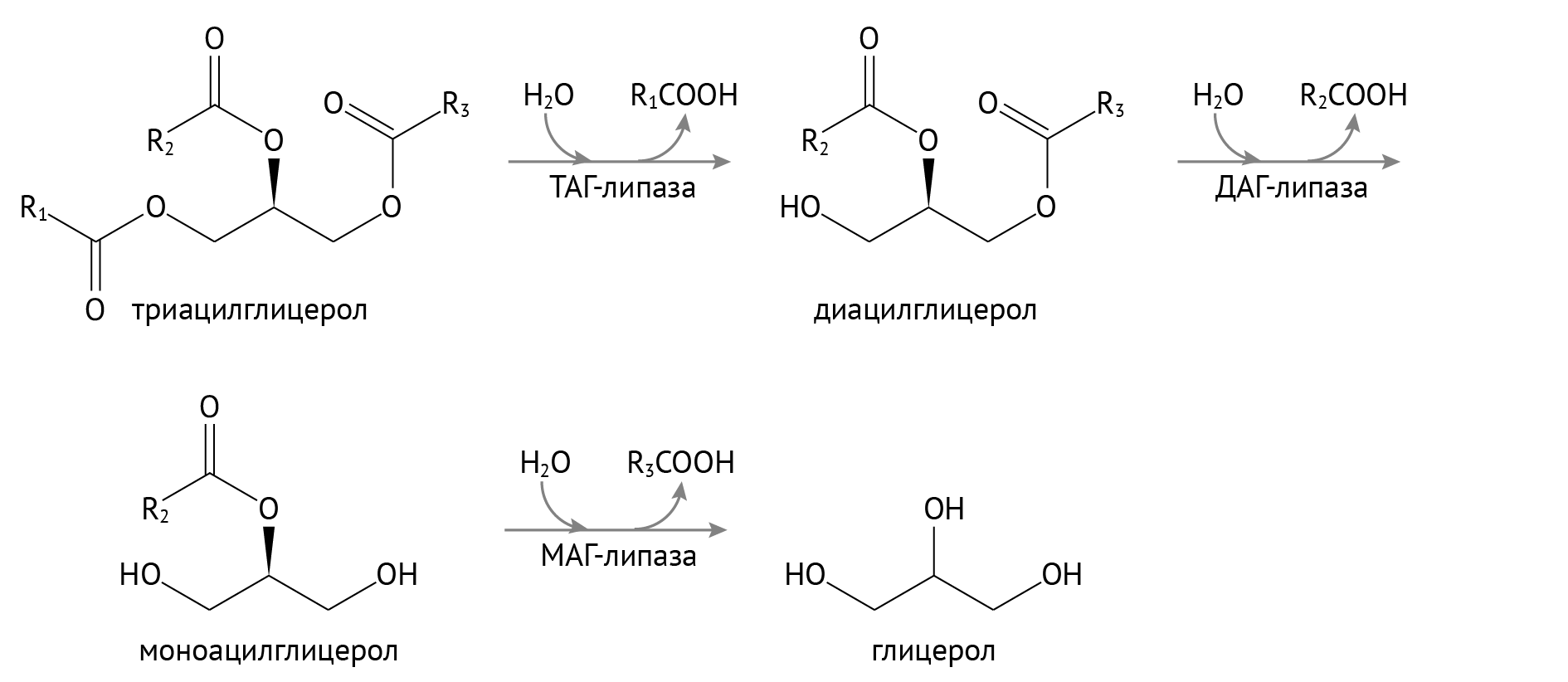
ТАГ-липаза является гормон-чувствительным ферментом, которая очень остро реагирует на критику и на действие контринсулярных гормонов — в первую очередь, адреналина и глюкагона. Он связан с активацией внутриклеточной протеинкиназы А. К слову, с активацией протеинкиназы А сопряжена и активация процесса распада гликогена в печени и мышечной ткани, идущая по аналогичному с активацией ТАГ-липазы механизму [3]:
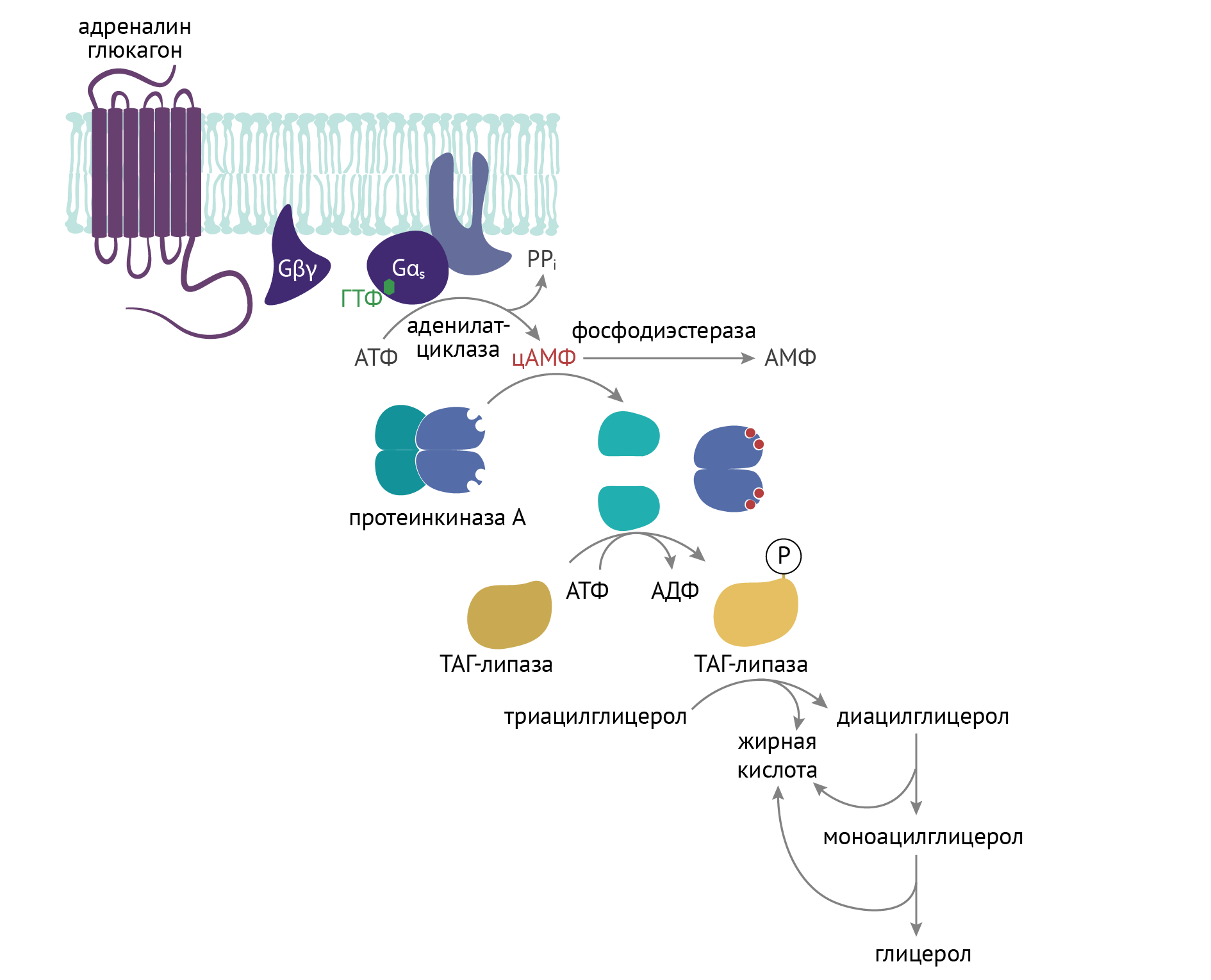
Не для зубрежки. Литературная минутка
* Большой и неуютный цитозоль. Здесь кипит жизнь, ничего не стоит на месте. Туда-сюда шныряют тРНК, унося с собой несчастную аминокислоту. А вот там можно заметить убиквитинлигазу, которая поймала нарушителя понятий и упаковывает его в протеасомы.
Здесь у каждого есть свое место. Это биологический мегаполис, в водоворот которого случайным образом (хотя что в нашей жизни вообще случайно?) попала жирная кислота. Одинокая, потерянная и несчастная. Слоняясь по клетке и зайдя не в тот угол, она встречается с ацил-КоА-синтетазой. Это встреча не проходит для нее бесследно. Теперь она жирная кислота, но с клеймом в виде S-KoA-группы, и способна вступать в неприличные связи с обитателями цитозоля клетки. Но ей хочется большего, и, как и в макромире, это не всегда приводит к добру…»
Ацил-КоА встречает на своем пути карнитин. Пообщавшись, жирная кислота узнает о существовании чрезвычайно увлекательного места под названием «митохондрия». Будучи легкомысленной и наивной, и в то же время чрезвычайно (в химическом отношении) активной, она недолго ломается и принимает предложение карнитина вписаться в эту удивительную семейную вечеринку.
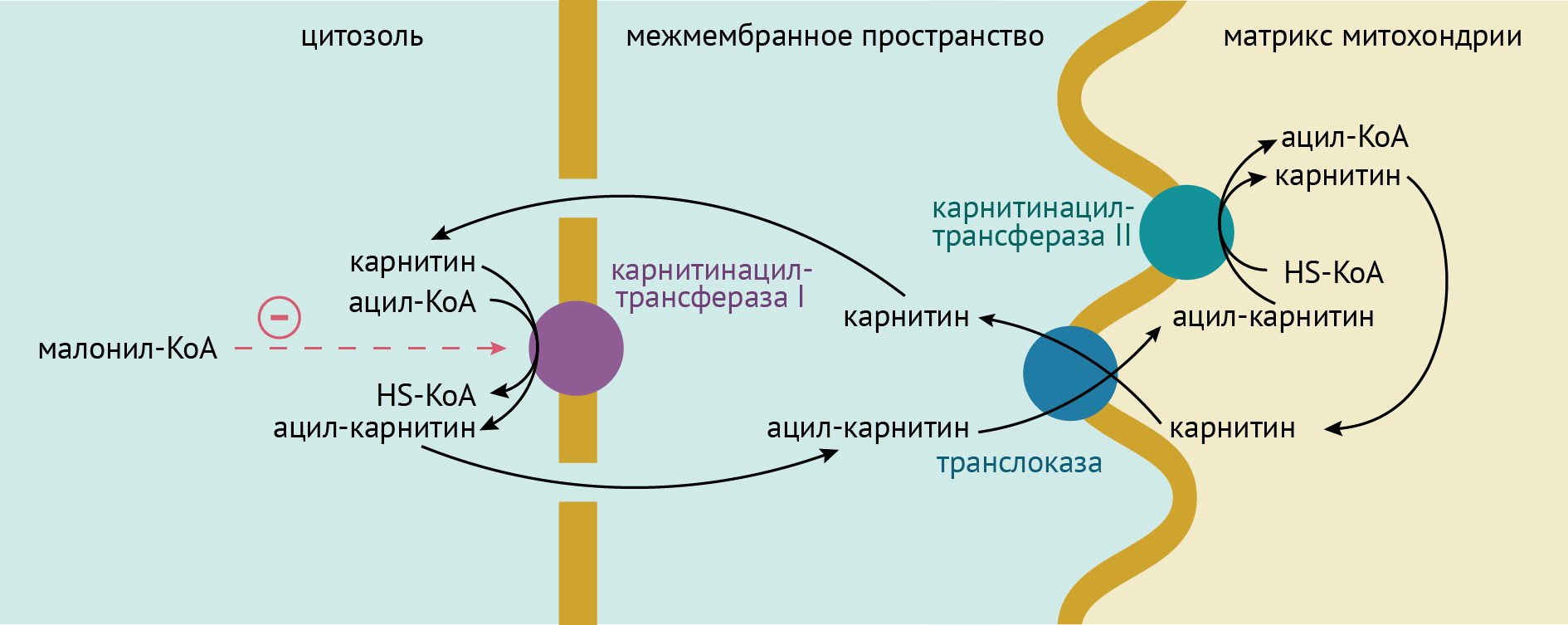
Карнитин — «свой» на этой вечеринке и проносит с собой жирную кислоту, кружась с ней в карнитинацилтрансферазном I танце. Достигая пика любви, карнитин увлекает жирную кислоту через транслоказу в митохондрию. Но на этом их ожидания начинают разниться и карнитин, оставив жирную кислоту с багажом воспоминаний (о котором стыдно рассказывать, но который приятно поворошить), уходит. Преодолев карнитинацилтрансферазу II, ацилкарнитин перестает существовать как пара и распадается на карнитин и ацил-КоА.
β-окисление жирных кислот
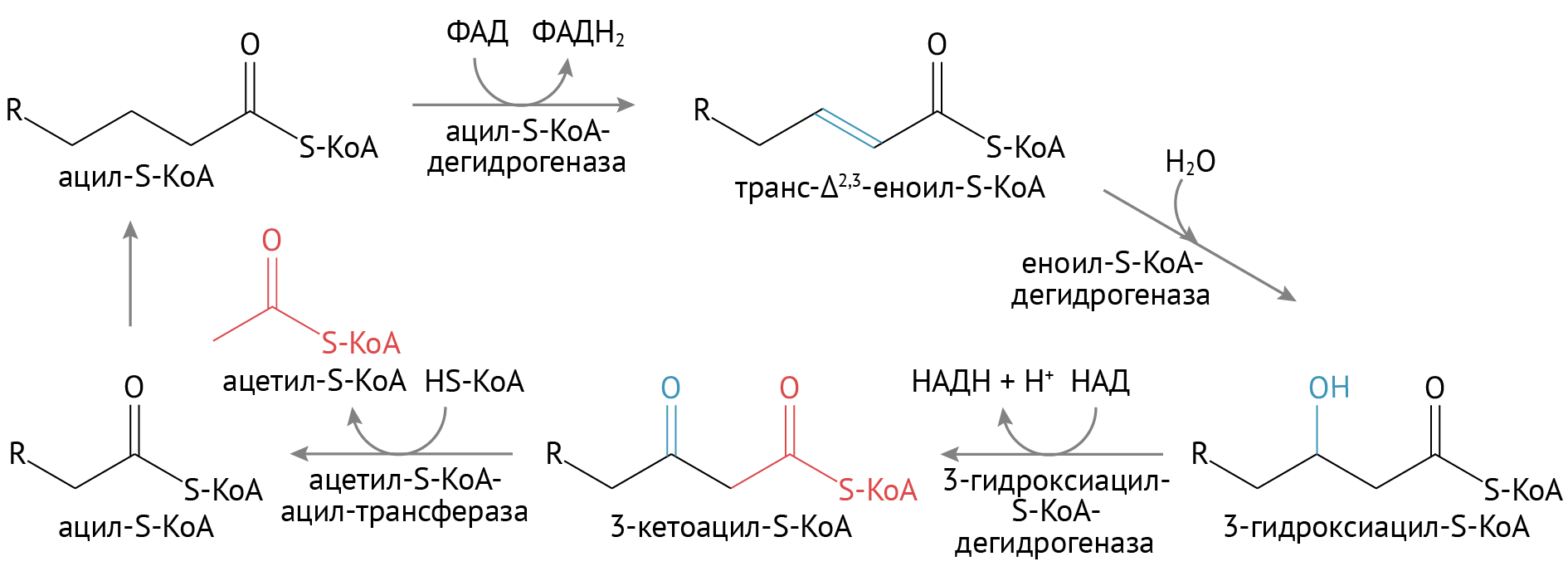
- Сверху представлена схема расщепления насыщенных жирных кислот, т. е. тех кислот, которые исходно не имеют в своей структуре двойных (ненасыщенных) ковалентых связей. Если мы имеем дело с исходно ненасыщенной кислотой, для нее первый цикл начнется со второй реакции.
- Конечный продукт — молекулы ацетил-КоА — необходимый субстрат для цикла трикарбоновых кислот.
- Получение энергии обусловлено восстановлением ФАД до ФАДН2 и НАД до НАДН + Н+. Эти восстановительные эквиваленты играют роль доноров электронов в электрон-транспортной цепи, и, в конечном счете, для получения энергии.
- В целом, окислением одной жирной кислоты можно получить в среднем 90–130 молекул АТФ. Это очень много, особенно для одной молекулы.
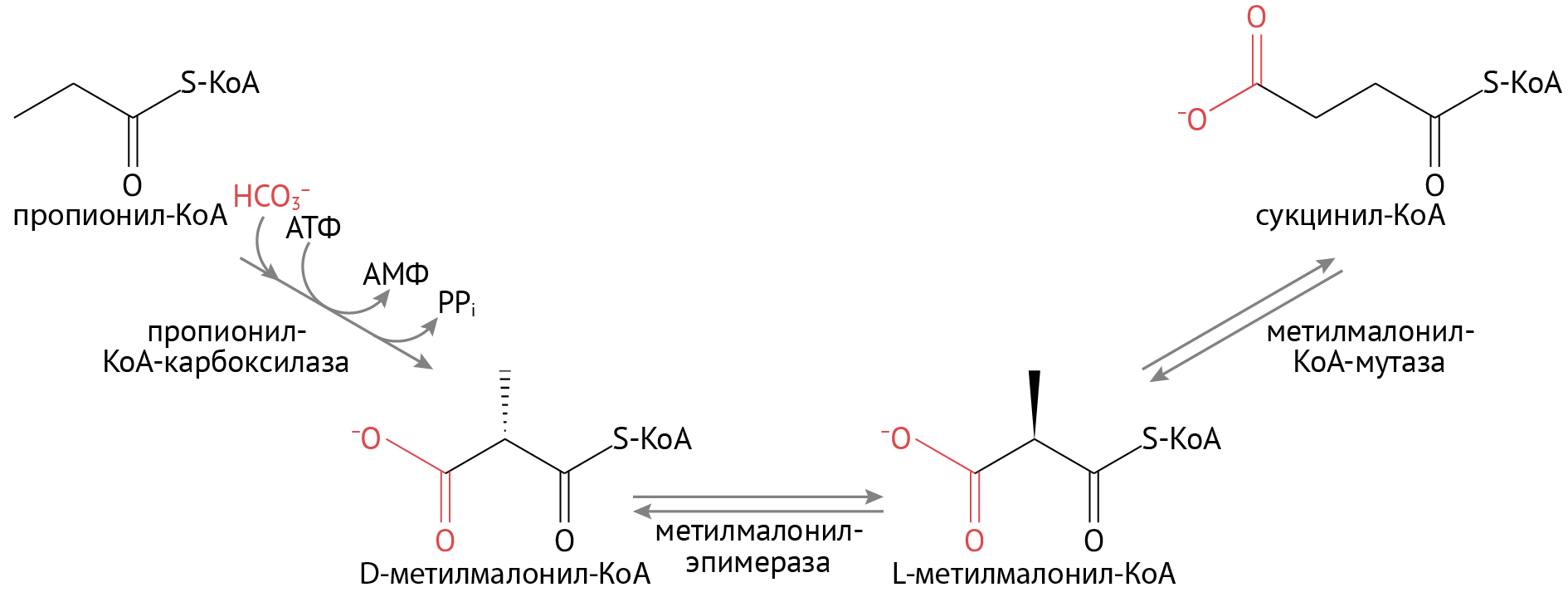
- На последней стадии окисления образуется не ацетил-КоА, а пропионил-КоА. Путем нехитрых превращений она станет сукцинил-КоА — активным участником того же ЦТК.
Читайте также: Плотная волокнистая соединительная ткань это внутренняя
Метаболизм углеводов и липидов характеризуется наличием еще и сложных анаболических реакций, которыми я пока не хочу нагромождать и без того огромнейшую главу. Какие-то моменты по анаболизму я поясню чуть позже, когда коснусь проблемы заболеваний.
Источники:
- Я. Кольман, К.-Г. Рём. Наглядная биохимия, 5-е издание, 2018 г., с. 154–168.
- Тимин О. А. Основы биологической химии, 2018 г., с. 232–257.
- Портал «Биохимия для студента»:, раздел «Липиды».
1. Катаболизм (распад) триацилглицеринов
Катаболизм триацилглицеринов можно разделить на 3 фазы:
1) гидролитическое расщепление 3-х эфирных связей;
2) катаболизм жирных кислот;
Гидролитическое расщепление триацилглицеринов
Гидролиз жиров является ступенчатым процессом (т.е. 3 эфирные связи в молекулах триацилглицеринов расщепляются не одновременно) и происходит при участии фермента липазы. Обнаружено несколько разновидностей липаз, содержащихся в клетках животных и растений. Липазы широко распространены в семенах и вегетативных органах растений. Скорость гидролиза сбалансирована со скоростью внутриклеточного окисления продуктов гидролиза: как правило, накопления жирных кислот и глицерина не происходит.
Процесс расщепления жира в растительном организме происходит особенно энергично при прорастании масличных семян. Он начинается с гидролитического распада жиров, происходящего под действием липазы и сопровождающегося накоплением глицерина и свободных жирных кислот. Образующиеся глицерин и жирные кислоты чрезвычайно быстро используются для различных процессов синтеза, протекающих в развивающемся ростке. При этом главным продуктом, возникающим в результате превращения жиров, является сахар. Установлено, что при прорастании богатых жиром семян образуются не только гексозы, но и пентозы. Этот факт указывает на то, что во время прорастания семени жиры расщепляются до низкомолекулярных соединений, содержащих два или три углеродных атома в молекуле. Путем конденсации этих низкомолекулярных соединений образуются затем различные моносахариды и другие вещества. Этот процесс происходит в особых субклеточных структурах – глиоксисомах и называется глюконеогенезом.
Частичное расщепление жиров пищи, особенно высокоэмульгированных (молоко, яичный желток) происходит в желудке, под действием липазы. Способность желудочной липазы расщеплять жир быстро уменьшается по мере увеличения длины цепей жирных кислот, входящих в состав жира.
В полости желудка действуют также желчь и ферменты поджелудочного сока, забрасываемые в желудок антиперистальтическими движениями главным образом при приеме жирной пищи, что обеспечивает более интенсивный гидролиз липидов.
Однако основное количество пищевых жиров расщепляется в 12-перстной кишке. В гидролизе жиров существенную роль играет желчь, выделяемая печенью. Известно, что пищевой жир, поступающий в тонкую кишку, в основном недоступен липазе из-за блокирования границы жировой массы белками или фосфолипидами. Желчные кислоты обладают способностью очищать границу эмульгированного жира от белка. В полости тонкой кишки под действием липазы образуются ди- и моноглицериды жирных кислот и незначительное количество свободного глицерина.
Катаболизм жирных кислот
Как известно, жиры отличаются от белков и углеводов своей высокой калорийностью: при окислении 1г нейтральных жиров выделяется около 9 ккал энергии (тогда как при окислении белков и углеводов только 4 ккал).
Почти половина энергии в процессах, протекающих в организме, извлекается из жирных кислот. При голодании жирные кислоты – единственный источник энергии.
Основным источником жирных кислот является гидролиз 3 ацилглицерола.
Читайте также: Ткани для шитья кукол барби
Важнейший этап их распада (диссимиляции) – так называемое β-окисление, открытое Ф. Кноопом.
При этом окисление жирной кислоты происходит у того углеродного атома, который находится в β-положении по отношению к карбоксильной группе.
При изучении прорастающих семян подсолнечника установлено, что ненасыщенные жирные кислоты легко превращаются в насыщенные. По-видимому, существенную роль в превращениях ненасыщенных жирных кислот играет липоксигеназа, катализирующая окисление этих кислот кислородом воздуха по месту локализации двойной связи.
Окисление предельных высших жирных кислот осуществляется ступенчато, путем отщепления от их молекул двууглеродных фрагментов. Все реакции многостадийного окисления ускоряются специфическими ферментами.
В результате происходит разрыв углеродной цепочки жирной кислоты между - и β-углеродными атомами с образованием ацетильного радикала и новой высокомолекулярной жирной кислоты, содержащей на 2 углеродных атома меньше, чем подвергшаяся окислению первоначальная жирная кислота.
Процесс β-окисления жирных кислот происходит в матриксе митохондрий при участии кофермента А и начинается с его присоединения к молекуле жирной кислоты (этот процесс является подготовительным, он обеспечивает активацию жирных кислот).
Процесс β-окисления жирных кислот осуществляется по спирали, в виде чередующихся циклов, каждый из которых состоит из 4-х реакций:
1) 1-е окисление: происходит отнятие водорода в -β-положении, осуществляемое под действием флавинового фермента;
2) гидратация: по месту двойной связи присоединяется молекула воды и образуется β-оксикислота;
3) 2-е окисление: образовавшаяся β-оксикислота подвергается окислению путем отнятия водорода, которое происходит при участии НАД+, что приводит к образованию β-кетокислоты
4) тиолиз: расщепление возникающей β-кетокислоты, происходящее под действием новой молекулы кофермента А:
В результате образуется ацетилкофермент А и соединенный с другим остатком коэнзима А радикал новой жирной кислоты, содержащей на 2 углеродных атома меньше, чем молекула исходной жирной кислоты. Новая кислота может снова подвергнуться β-окислению, пока не окислится полностью. Все реакции идут в матриксе митохондрий.
Таким образом, β-окисление представляет из себя окисление жирных кислот в β-положении и превращение их в β-кетокислоты, которые расщепляются с образованием остатка уксусной кислоты и жирной кислоты, укороченной на 2 атома углерода.
Конечным продуктом β-окисления является ацетил-КоА. Он либо полностью окисляется до СО2 и Н2О, вступая в цикл Кребса, либо используется на синтез углеводов (у растений) и новых жирных кислот.
Продуктом окисления жирных кислот с нечетным числом атомов углерода, наряду с ацетил-КоА, является пропионил-КоА, который превращается в сукцинил-КоА, последний вовлекается в цикл Кребса.
ФАДН2 и НАДН, образующиеся при β-окислении, окисляются в дыхательной цепи с одновременным образованием пяти молекул АТФ. Поскольку на 1-м этапе β-окисления расходуется 1 молекула АТФ, то «чистый» выход АТФ на 1-м этапе составляет 4 молекулы АТФ.
Таким образом, при полном окислении до углекислого газа и воды одной молекулы пальмитиновой кислоты С16Н32О2 выход продуктов реакции и АТФ будет следующий.
В результате семи этапов β-окисления, на каждом из которых образуется 5 молекул АТФ, возникает 35 молекул АТФ. С вычетом одной молекулы АТФ, расходуемой на 1-м этапе, получаем 34 молекулы.
Полное окисление восьми молекул ацетил-КоА в цикле Кребса дает 96 молекул АТФ (окисление одной молекулы ацетил-КоА дает 12 молекул АТФ).
В общей сложности образуется 16 молекул СО2 и 16 молекул Н2О и возникает 130 молекул АТФ.
Наряду с β-окислением, высокомолекулярные жирные кислоты (С14,С16, С18) в некоторых растительных тканях могут подвергаться также -окислению. При этом процесс начинается с декарбоксилирования жирной кислоты, которое происходит под действием особой пероксидазы и пероксида водорода. В результате этого этапа образуется соответствующий альдегид, содержащий на 1 углеродный атом меньше, чем исходная жирная кислота. Альдегид далее подвергается окислению под действием особого фермента альдегид-дегидрогеназы, коферментом которого является НАД. Синтезируется жирная кислота, содержащая на 1 углеродный атом меньше, чем исходная. Эта жирная кислота может снова подвергнуться описанному выше декарбоксилированию с последующим окислением образовавшегося альдегида. По-видимому, физиологическая роль процесса -окисления заключается в образовании жирных кислот с нечетным числом атомов углерода.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом