
Большинство генов белков промежуточных филаментов у человека кодируют кератины. Как показано в таблицах ниже, известно 28 кератинов, относящихся к типу I, и 26 белков, относящихся ко II типу. В ткани эпителия кератины, которые иногда называются цитокератинами, экспрессируются совместно, т. е. как тип I/тип II. Экспрессия кератинов является настолько характерной особенностью клеток эпителия, что их присутствие может служить признаком эпителиальной ткани. Ткани, которые не экспрессируют кератины (например, клетки эндотелия кровеносных сосудов) не относятся к эпителиальным.
Кератиновые филаменты связаны с межклеточными контактами (десмосомами) и с местами контактов клетки с матриксом (полудесмосомами).
Все вместе образует структурную сеть, которая особенно характерна для многослойного эпителия, например эпидермиса. Основные кератины эпителиального слоя подразделяются на группы в зависисмости от заряда и молекулярной массы. В состав группы К1 (тип II) входят наиболее крупные и основные белки, а в состав группы К19 — более мелкие и кислые белки группы I.
Каждый кератин группы I экспрессируется совместно с кератином группы II, и каждая пара характерна для степени дифференцировки и специализации определенного типа клеток эпителия. Хотя in vitro любой из кератинов, относящихся к I или II типу, будет образовывать филаменты с различными комплементарными кератинами другого типа, in vivo они проявляют гораздо большую селективность, образуя специфические пары.
Экспрессия таких пар кератинов тесно связана со специфическими направлениями дифференцировки эпителия или даже с различными стадиями этого процесса, и присутствие одного представителя пары почти всегда служит признаком наличия другого. С функциональной точки зрения, пары кератинов можно подразделить по крайней мере на три группы: простые, барьерные и структурные кератины.

Экспрессия кератинов характеризуется тканевой специфичностью.
Они экспрессируется попарно: тип I/тип II.
Каждая пара характерна для специфического типа дифференцировки эпителия. Выделены первичные кератины.
Кератины экспрессируются в клетках эпителия. Эпителиальная ткань представляет собой тип тканевой организации, который характеризуется плотной упаковкой клеток в один или несколько слоев. Эпителий образует границу органов, а также секреторных и всасывающих протоков. В этих случаях обычно говорят об однослойном эпителии, клетки которого непосредственно контактируют с подлежащим слоем внеклеточного матрикса (базальной ламины), а свободная поверхность открыта в просвет протока или кишки.
Крайний случай представляет собой многослойный или стратифицированный эпителий, образующий основную барьерную ткань оболочек тела и органов. К этому типу относятся эпидермис, покрывающий внешнюю поверхность тела, а также специализированный эпителий, выстилающий отверстия и полости прилегающих к ним протоков. Клетки стратифицированного эпителия обычно называют кератиноцитами. Ткань стратифицированного эпителия также формирует дополнительные структуры эмбрионального происхождения, состоящие из специализированного сложного эпителия, образующего, например, железы, волосы и ногти, которые могут включать участки, состоящие из простого эпителия.
Наружные клетки полностью сформировавшегося стратифицированного барьерного эпителия плоской или чешуйчатой формы. Этот эпителий обычно состоит из 6-10 слоев клеток Чтобы защитить тело от физических, химических и канцерогенных воздействий, клетки постоянно обновляются.
Клетки эпителия скрепляются вместе посредством десмосом, а также полудесмосом с субстратами. Все контакты в виде плотной сети, состоящей из связок кератиновых филаментов, проходят через цитоплазму каждой клетки. По сравнению с клетками других типов, кератиноциты многослойного эпителия экспрессируют большее количество и более широкий набор белков промежуточных филаментов, что связано с тем, что эти клетки должны противостоять массивным внешним физическим воздействиям.
Состав специфических белков кератиновых филаментов, которые экспрессируются в эпителиальных клетках, зависит от локализации и физиологического состояния последних. Особенно отчетливо это прослеживается в изменении экспрессии кератина, которое происходит при дифференцировке клеток стратифицированного эпителия. К числу наименее дифференцированных клеток стратифицированного эпителия принадлежат базальные клетки, которые еще сохраняют контакт с базальной мембраной. Эти клетки еще способны к делению, некоторые из них являются стволовыми клетками ткани, и делиться крайне редко, однако большая часть клеток делится, обеспечивая тем самым рост ткани.
После того как базальная клетка разделилась, одна из дочерних клеток покидает базальный компартмент и перемещается в первый супрабазальный слой. В результате этого важного перемещения клетка выходит из-под прямого влияния базальной мембраны и сигналов роста и становится коммитированной к терминальной дифференцировке. Затем клетка начинает свой путь к поверхности эпителия. Этот путь представляет собой процесс терминальной дифференцировки, который в конце концов завершается гибелью клетки и ее потерей из ткани. Рисунок ниже иллюстрирует прогрессирующие изменения экспрессии кератинов, связанные с выходом некоторых клеток из пролиферативного компартмента и их коммитированием к дифференцировке.
Читайте также: Как сделать мягкий шарик из ткани для кота
В процессе развития первыми экспрессируются простые кератины. В этой группе первичные или универсальные кератины представлены белками К8 (тип II) и К18 (тип I). Они присутствуют в ранних эмбриональных клетках и, по-видимому, представляют собой эволюционно наиболее древние кератины. Их экспрессия минимальна, что соответствует наименее дифференцированному состоянию кератинов, характерному для функционального эпителия, пласт которого состоит из поляризованных и плотно, без промежутков, скрепленных между собой клеток.
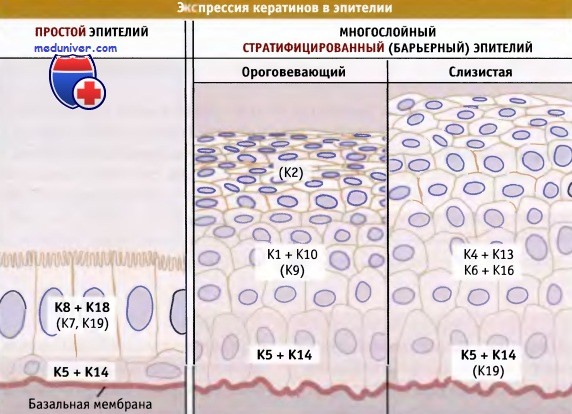
Последовательность экспрессии основных кератинов в однослойном (например, в железистом) и барьерном (например, в ороговевающем или слизистом) эпителии.
Первичные кератины выделены жирным шрифтом. Вариабельные или минорные кератины не показаны.
Экспрессия кератинов зависит от положения клеток в ткани и, таким образом, от ее пролифертивного статуса:
утрата контакта клеток с базальным слоем приводит к их выходу из цикла и к вступлению в дифференцировку.
У позвоночных К8 и К18 представляют собой белки с наиболее консервативной первичной структурой и присутствуют в клетках на всех ступенях развития, от яйцеклетки до зрелой ткани. Все эмбриональные клетки экспрессируют К8 и К18 до стадии гаструляции, когда некоторые эктодермальные клетки начинают дифференцироваться, образуя слой мезодермы, прекращают синтез К8/К18, и начинают экспрессировать виментин, белок, относящийся к типу III.
Белки К8 и К18 продолжают экспрессироваться в клетках эмбрионального эпителия до тех пор, пока эти клетки не станут коммитированными к специфическому направлению морфогенеза, т. е. когда они начинают экспрессировать тканеспецифические белки промежуточных филаментов. Во взрослом организме К8 и К18 характерны для клеток простого эпителия, обладающих секреторными и/или всасывающими функциями, например клеток желез, печени, дыхательного эпителия и желудочно-кишечного тракта. Эти белки экспрессируются в клетках различных карцином, и антитела к ним широко используются для диагностики различных патологических состояний. Наряду с первичными кератинами, известны, по меньшей мере, еще два простых кератина, К7 (близкий к К8 в основном содержится в клетках протоков желез) и К20 (близкий к К18, который содержится в отделах желудочно-кишечного тракта).
Барьерные кератины характерны для стратифицированного эпителия. К первичным, или основным, кератинам этой группы относятся К14 (тип I) и К5 (тип II), присутствующие в кератиноцитах базального слоя стратифицированного сквамозного эпителия, например эпителия кожи. Этот слой, состоящий из одного ряда базальных клеток, экспрессирующих К5/К14, представляет собой наименее дифференцированный тканевой компартмент, сохраняющий способность к пролиферации. В сложном железистом эпителии, наряду с простыми клетками, экспрессирующими К8/К18, находятся базальные клетки, экспрессирующие К5/К14. В некоторых тканях базальные пролиферирующие клетки экспрессируют другие кератины, например К19, К15 и К6/К17.
Когда клетки выходят из базального слоя, они прекращают продуцировать К5/К14 и начинают экспрессировать вторую пару кератинов, специфичную для дифференцирующихся клеток. В эпидермисе это кератины К1 типа II и К10 типа I. Экспресия вторичных кератинов в супрабазальных слоях клеток зависит от типа ткани. Один набор кератинов, представляющий собой изоформы К6, а также К16 и К17, проявляет свойства стрессорных белков эпидермиса, и их синтез индуцируется вскоре после раневых повреждений и воспалительного процесса. В других тканях эти «стрессорные» кератины экспрессируются постоянно, и этим тканям свойственна роль характерных индукторов воспаления.
Таким образом, пролиферативный компартмент (базальный слой клеток) многослойного барьерного эпителия характеризуется спектром синтеза кератинов отличным от дифференцирующегося компартмента (супрабазальных слоев). Для этих барьерных тканей выход клеток из цикла тесно связан с остановкой синтеза первичных кератинов и с началом синтеза вторичных, специфичных для процесса дифференцировки. Кератины клеток супрабазального слоя обеспечивают большую тканевую упругость, и экспрессия нескольких вторичных кератинов может препятствовать делению клеток в митозе и, таким образом, несовместима с пролиферацией.
Третья группа «структурных» кератинов включает большое количество белков промежуточных филаментов, которые экспрессируются только в специализированных поддерживающих структурах, таких как волосы и ногти. К ним относятся кератины клеток волос или трихоцитов, образующих эти структуры, а также специализированные кератины, образующиеся в продуцирующих их эпителиальных клетках. Первичная структура этих двух типов структурных кератинов различна, и считается, что у позвоночных они представляют собой поздний продукт эволюции.
Читайте также: Ткань ункл 3 характеристики
Последовательная экспрессия структурных кератинов особенно характерна для волосяных фолликулов. Первая группа структурных кератинов экспрессируется в концентрических слоях эпителиальных трубочек внутреннего корневого чехла волосяного фолликула. Эта структура становится очень прочной и формирует основную часть волоса. Вторая группа опорных кератинов (трихоцитов или клеток волоса) экспрессируется в основной части волоса, ногтях, некоторых опорных клетках поверхностных сосочков языка (что характерно для языка кошачьих), и в небольшом количестве в тимусе. Эти кератины трихоцитов отличаются высоким содержанием цистеина и пролина в головных и хвостовых доменах.
Последнее позволяет им образовывать дисульфидные связи с белками, которые находятся в цитоплазме клеток волоса по мере дифференцировки и упрочения последних. Эти белки не входят в состав филаментарных структур и называются белками, связывающимися с кератином. Наличие дисульфидных связей приводит к образованию очень прочных структур, формирующих поддерживающую ткань.
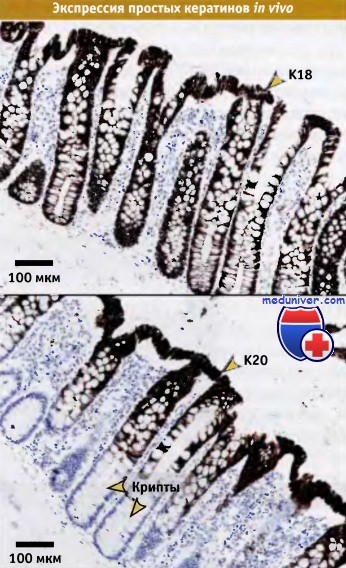 Иммуногистохимическое окрашивание простых кератинов на срезе слизистой толстого кишечника.
Иммуногистохимическое окрашивание простых кератинов на срезе слизистой толстого кишечника.
Локализация двух кератинов I типа, К18 и К20 определена с помощью двух моноклональных антител с последующим иммунопероксидазным выявлением.
Кератины окрашиваются коричневым цветом на фоне голубой окраски гематоксилином.
Первичный кератин К18 находится во всех клетках простого эпителия, в то время как К20 специфичен для определенных клеток желудочно-кишечного тракта.
Как видно на препарате, его присутствие отмечено только в более дифференцированных клетках эпителия, по мере миграции клеток вверх от основания крипты. 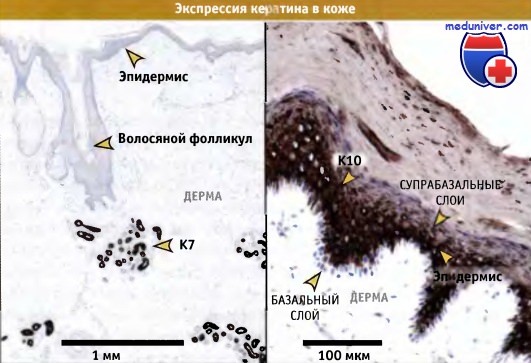 Тканевая специфичность экспрессии кератина в коже.
Тканевая специфичность экспрессии кератина в коже.
На срезах, окрашенных антителами к К7, простому кератину типа II, видны только секреторные клетки потовых желез (иммунопероксидазой окрашиваются в темно-коричневыц цвет).
Многослойный ороговевающий барьерный эпителий (эпидермис) не окрашивается антителами, и после окраски гематоксилином ядра имеют бледно-голубую окраску.
Справа: срез плотного эпидермиса, окрашенный антителами на К10,
тип I вторичный или тканеспецифичный кератин, экспрессирующийся в постмитотических супрабазальных клетках ороговевших барьерных тканей.
Базальный слой клеток не окрашивается антителами, и ядра окрашены в синий цвет. 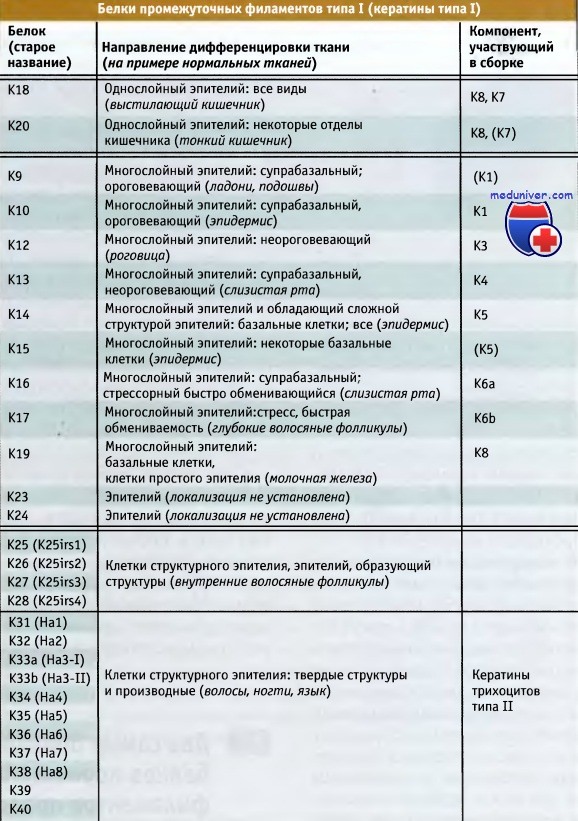 Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа I (кератины типа I).
Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа I (кератины типа I).
Белки сгруппированы в простые кератины (клетки простого эпителия), барьерные кератины (характерные для многослойного чешуйчатого эпителия и эпителия сложной структуры)
и в два типа структурных кератинов, связанных с эпидермальными элементами  Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа II (кератины типа II).
Белки промежуточных филаментов человека, относящиеся к гомологичной группе типа II (кератины типа II).
Белки сгруппированы в простые кератины (клетки простого эпителия),
барьерные кератины (характерные для многослойного ороговевающего эпителия и эпителия сложной структуры)
и в два типа структурных кератинов, связанных с эпидермальным элементами.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Кератиновые филаменты в эпителиальной ткани
Кафедра патологической анатомии медицинского факультета ФГБОУ ВПО «Российский университет дружбы народов»
Отделение хирургической стоматологии ФГБУ «Центральный научно-исследовательский институт стоматологии и челюстно-лицевой хирургии» Минздрава РФ, Москва
кафедра патологической анатомии Российского университета дружбы народов, Москва, Россия, 117198
Цитокератин 15 как диагностический маркер начала малигнизации эпителия слизистой оболочки рта
Журнал: Стоматология. 2018;97(6): 61-62
Ивина А. А., Семкин В. А., Бабиченко И. И. Цитокератин 15 как диагностический маркер начала малигнизации эпителия слизистой оболочки рта. Стоматология. 2018;97(6):61-62.
Ivina A A, Semkin V A, Babichenko I I. Cytokeratin 15 as a diagnostic marker for oral epithelial malignization. Stomatologiya. 2018;97(6):61-62. (In Russ.).
https://doi.org/10.17116/stomat20189706161
Кафедра патологической анатомии медицинского факультета ФГБОУ ВПО «Российский университет дружбы народов»



В работе представлен пример иммуногистохимического исследования случая малигнизации эпителия при очаговой эпителиальной гиперплазии слизистой оболочки рта (СОР). В описанном случае ввиду сложности определения начала малигнизации эпителия при обычной гистологической методике был применен иммуногистохимический метод с антителами к белкам Ki-67, вирусу папилломы человека 16-го типа и цитокератину 15. В результате иммуногистохимического исследования установлены повышенная пролиферативная активность эпителиальных клеток, наличие в их цитоплазме папилломавируса HPV16 и синтез цитокератина 15 в клетках всех слоев эпителия, что характерно для плоскоклеточной интраэпителиальной неоплазии и может быть диагностическим критерием начала малигнизации эпителия СОР.
Кафедра патологической анатомии медицинского факультета ФГБОУ ВПО «Российский университет дружбы народов»
Отделение хирургической стоматологии ФГБУ «Центральный научно-исследовательский институт стоматологии и челюстно-лицевой хирургии» Минздрава РФ, Москва
кафедра патологической анатомии Российского университета дружбы народов, Москва, Россия, 117198
Одним из актуальных направлений современной стоматологии является ранняя диагностика предраковых состояний слизистой оболочки рта (СОР). Это необходимо для профилактики развития злокачественных опухолей, среди которых по частоте встречаемости 1-е место занимает плоскоклеточный рак (ПР). На его долю приходится более 90% злокачественных новообразований СОР. Пятилетняя выживаемость при диагностике ПР на поздней стадии невысока — около 20% [1]. Известно, что вирус папилломы человека 16-го типа (ВПЧ16) лежит в основе малигнизации эпителия [2]. Поэтому выявление злокачественного перерождения эпителиальных клеток СОР на фоне папилломавирусной инфекции позволит своевременно провести лечение и улучшить качество и продолжительность жизни пациентов.
Читайте также: Название ткани для юбки для школы
При гистологическом исследовании на срезах, окрашенных гематоксилином и эозином, не всегда удается точно выявить начало малигнизации эпителиальных клеток СОР. Особенно сложно это сделать на фоне выраженного хронического воспаления в собственной пластинке СОР. Плоскоклеточная интраэпителиальная неоплазия I—II степени (squamous intraepithelial neoplasia — SIN1—2) является первым признаком злокачественных изменений эпителия СОР. Постановка данного диагноза с обнаружением ВПЧ16 во многом определяет тактику дальнейшего ведения пациента. Учитывая актуальность проблемы и сложность диагностики SIN1—2, приводим наблюдение из практики.
Пациентка К., 55 лет, обратилась в отделение хирургической стоматологии ЦНИИС и ЧЛХ Минздрава России с жалобами на ощущение дискомфорта и локального уплотнения языка справа. При осмотре на боковой поверхности языка справа определялся участок лейкоплакии. Проведено иссечение патологического очага.

При гистологическом исследовании выявлены гиперплазия базального и шиповатого слоев многослойного плоского эпителия, явления пара- и гиперкератоза и выраженная хроническая воспалительная реакция в собственной пластинке СОР (см. рисунок, Гиперпластические и неопластические изменения эпителия СОР. ОЭГ: а—г, плоскоклеточная интраэпителиальная неоплазия — д—з (а, д — гематоксилин и эозин; б, е — ИГХ-реакция с маркером Ki-67; в, ж — ИГХ-реакция с маркером ВПЧ16; г, з — ИГХ-реакция с маркером СК15; ИГХ окрашивание — ДАБ—гематоксилин Майера; ув. 100). а). На одном из участков отмечались активно пролиферирующие эпителиальные клетки базального и парабазального слоев с умеренной ядерной атипией, а под базальной мембраной — хронический воспалительный инфильтрат (см. рисунок, д).
Гистологическая картина соответствовала очаговой эпителиальной гиперплазии (ОЭГ), однако было решено провести иммуногистохимическое (ИГХ) исследование с целью уточнения диагноза и исключения начала малигнизации эпителия СОР, так как активно пролиферирующие клетки и умеренная ядерная атипия могут возникнуть как реакция на хронический воспалительный процесс в собственной пластинке, а также и в результате злокачественной трансформации эпителиоцитов СОР.
ИГХ-исследование биопсийного материала проводилось в соответствии со стандартным протоколом. Тканевые антигены определяли с помощью мышиных моноклональных антител к Ki-67 (ММ1, Diagnostic Biosystems), поликлональных кроличьих антител к ВПЧ16 (ABBIOTEC) и мышиных моноклональных антител к цитокератину 15 (СК15) (ЕР14, Epitomics). Иммуногистохимическое исследование осуществлялось в автоматическом режиме на Autostainer 360 (Thermo scientific) с использованием системы визуализации QUANTO.
При ИГХ-исследовании установлено незначительное повышение пролиферативной активности эпителиальных клеток парабазального слоя, что соответствовало ОЭГ (см. рисунок, б). В участке, подозрительном по малигнизации, пролиферирующие клетки отмечались не только в базальном и парабазальном слоях, но и в шиповатом; была видна также умеренная ядерная атипия, что соответствует диспластическим изменениям ткани (см. рисунок, е).
Антигены ВПЧ высокого онкогенного риска — ВПЧ16 при ОЭГ не выявлены (см. рисунок, в), а в участке дисплазии цитоплазма кератиноцитов парабазального и шиповатого слоев содержала этот вирус (см. рисунок, ж).
В отличие от ОЭГ, при которой белок СК15 выявляется только в клетках базального слоя (см. рисунок, г), при дисплазии отмечался выраженный синтез этого белка в цитоплазме клеток базального и шиповатого слоев, где ранее были установлены повышенная пролиферация и наличие ВПЧ16 (см. рисунок, з).
Результаты ИГХ-исследования соответствуют характеристикам SIN2 эпителия СОР, когда в процессе злокачественной трансформации клеток увеличивается их пролиферативная активность [3]. Это может быть связано с распределением ВПЧ высокого онкогенного риска — ВПЧ16 [2].
Цитокератины (кератины) — белки, которые формируют промежуточные филаменты в эпителиальных клетках. Вместе с актиновыми микрофиламентами и микротрубочками промежуточные филаменты выстраивают цитоскелет эукариотической клетки. Описано 20 разных кератинов (К1—К20), которые подразделяются на кислые кератины 1-го типа (9—20) и основные кератины 2-го типа (1—8). Кератиновые филаменты группируются как облигатные гетерополимеры, состоящие из мономеров 1-го и 2-го типов [4]. Белок СК15 относится к 1-му типу кератинов, который не имеет белка-партнера 2-го типа. Недавние исследования показали, что в неизмененном эпителии синтез К15 отмечается только в клетках базального слоя [5]. Появление СК15 в цитоплазме клеток выше базального слоя при SIN, вероятнее всего, связано с тем, что пролиферирующие клетки, которые в норме локализуются только в базальном слое, перемещаются вверх без должной дифференцировки, что характерно для злокачественной трансформации.
Таким образом, синтез белка СК15 в цитоплазме клеток парабазального и вышележащих слоев может служить диагностическим критерием начала малигнизации эпителия СОР.
Авторы заявляют об отсутствии конфликта интересов.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом