
Области применения метода культуры клеток и тканей.
Метод культуры тканей широко используется в сельском хозяйстве и промышленном производстве (рис. 8). Примером может служить массовое клональное микроразмножение плодовоовощных и декоративных растений, а также их оздоровление от вирусных и других инфекций. С помощью культуры in vitro можно расширить возможности селекционной работы: получать клоны клеток, а затем и растения с запрограммированными свойствами. Благодаря способности клеток синтезировать в культуре вторичные метаболиты возникла отрасль промышленности, осуществляющая биологический синтез веществ, необходимых человеку.
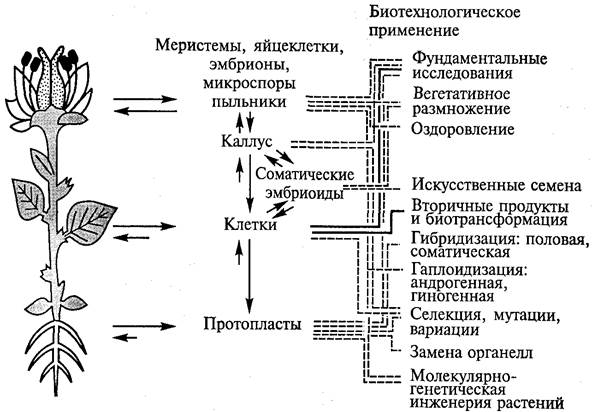
Рис. 8. Использование культуры клеток и тканей растений в биотехнологии (по Х. Борнман, 1991)
В настоящее время известно примерно 2 × 10 4 синтезируемых растениями веществ, которые используются человеком, и их количество постоянно увеличивается. Растения всегда служили источником пищи, эфирных масел, красителей и, конечно же, лекарственных соединений. Так, мак снотворный (Рараvеr sоттnifеrит) является источником болеутоляющего вещества — кодеина; из наперстянки (Digitalis lапаtа) получают дигоксин, тонизирующий сердечную деятельность; из хинного дерева (Сinсhопа ledgеriапа) — антималярийное средство «хинидин».
Особое место занимают наркотики и стимулирующие вещества. В небольших, строго контролируемых количествах их используют в медицине. Однако при систематическом употреблении низких концентраций наркотиков возникают наркозависимость и стремление к увеличению употребляемой дозы. Применение высоких концентраций наркотика убивает человека. Наиболее известны опиум и героин из Рараvеr sоттnifеrит, кокаин из Erythroxylon, никотин из различных сортов табака. Наиболее известный стимулятор — кофеин, содержащийся в растениях чая и кофе. Стимуляторы не токсичны в концентрациях, рекомендуемых к применению, однако высокие их концентрации негативно влияют на сердечно-сосудистую и нервную систему человека.
Большой интерес вызвало открытие пиретринов, выделенных из цветков Сhrуsапthemum cinerariaefolium. Эти вещества — мощные инсектициды. Особая их ценность заключается в том, что пиретрины не вызывают привыкания у насекомых, а также не проявляют кумулятивного токсического эффекта.
Способность растений синтезировать различные соединения привела к предположению, что тем же свойством будут обладать клетки и ткани этих растений, выращиваемые в стерильных условиях. Для некоторых культур это оказалось справедливым, но в отдельных случаях клетки либо не проявляли способности к синтезу необходимых веществ, либо синтезировали их в минимальных количествах. Понадобились долгие эксперименты по подбору питательных сред, условий культивирования, исследованию новых штаммов, полученных благодаря генетической гетерогенности каллусных клеток или применению мутагенных факторов, чтобы добиться серьезных успехов в этой области.
В настоящее время промышленный синтез вторичных метаболитов — очень перспективное направление. Синтез вторичных метаболитов происходит главным образом в суспензионной культуре клеток, в регулируемых условиях, поэтому он не зависит от климатических факторов, от повреждения насекомыми. Культуры выращивают на малых площадях в отличие от больших массивов плантаций с необходимыми растениями. Культуры клеток растений могут синтезировать практически все классы соединений вторичного обмена, причем довольно часто в количествах, в несколько раз превышающих их синтез в целых растениях. Например, выход аймалицина и серпентина в культуре клеток Саtharanthus roseus составляет 1,3% сухой массы, а в целом растении — 0,26%. В культуре клеток Dioscorea deltoidea диосгенин синтезируется в количестве 26 мг на 1 г сухой массы, а в клубнях растений его содержание составляет 20 мг на 1 г сухой массы.
Читайте также: Ткань pura lana vergine
Кроме того, в культурах клеток может начаться синтез веществ, не характерных для исходного растения, либо расширяется набор синтезируемых соединений. В ряде случаев в клеточной культуре образуются вещества, которые синтезировались интактным растением на ювенильной фазе развития, либо вещества, содержавшиеся в клетках филогенетически более ранних групп растений. Так, в культуре клеток Рараvеr bracteatum содержится сангвирин, характерный для ювенильных растений, и отсутствует тебаин, синтезируемый взрослыми растениями.
Важная особенность культивируемой популяции клеток — ее стабильность в отношении синтеза и накопления продуктов вторичного синтеза. Так, российскими учеными были получены разные штаммы клеток Dioscorеа deltoidea, в том числе штамм-сверхпродуцент ИФР ДМ-0,5. Все эти штаммы сохраняли стабильность в отношении синтеза фуростаноловых гликозидов около 26 лет.
Интересная особенность большинства клеток в культуре состоит в том, что обычно эти клетки не транспортируют синтезируемые метаболиты в питательную среду или другие клетки, хотя некоторые культуры составляют исключение, в частности культура клеток мака, которые депонируют алкалоиды в млечники. Синтез вторичных метаболитов в культивируемых клетках связан с внутриклеточными органеллами, в основном с пластидами и эндоплазматическим ретикулумом. В клетках, не способных к транспорту метаболитов, продукты вторичного синтеза обычно накапливаются в вакуолях и свободном пространстве клеток.
На синтез вторичных метаболитов влияет целый ряд факторов. Прежде всего, выход продукта зависит от генотипа растения-донора. Показано, что культуры клеток, полученных от высокопродуктивных растений, продуцировали большее число метаболитов. Другой важный фактор — состав питательной среды и концентрация ее компонентов, которые должны обеспечивать, с одной стороны, увеличение количества клеток-продуцентов, с другой — усиливать сам процесс синтеза. На рост, т.е. на увеличение биомассы, существенно влияет природа и количество углеводов, соединений азота и фосфора, на синтез метаболитов — природа и концентрация фитогормонов. Так, при замене одного ауксина на другой, например нафтилуксусной кислоты на 2,4-D (2,4-дихлорфеноксиуксусную кислоту), трехкратно увеличился синтез антрахинона суспензионной культурой Morinda citrifolia.
Существует современная технология получения вторичных метаболитов с помощью иммобилизованных клеток культуры, т. е. помещение их в определенный носитель или адсорбция в нем. Носитель с клетками помещают в питательную среду. Клетки остаются живыми. Они прекращают рост, но продолжают синтез метаболитов, выделяя их в среду.
Еще один из примеров использования вторичных метаболитов растений — получение карденолидов, гликозиды которых используют в медицине для лечения болезней сердца. Растения наперстянки (Digitalis lanata) в большом количестве синтезируют дигитоксин вместо необходимого дигоксина. Для соответствующей биотрансформации с успехом используют недифференцированную суспензионную культуру наперстянки. Иммобилизованные клетки этой культуры способны долгое время с постоянной скоростью трансформировать β-метил-дигитоксин в β–метилдигоксин.
Таким образом, использование суспензионных культур для синтеза вторичных метаболитов в промышленных масштабах имеет большие перспективы, и не только с точки зрения экономической выгоды получения более дешевой продукции в запланированных количествах. Важно, что использование культуры клеток может спасти от уничтожения тысячи дикорастущих растений, ставших уже редкими, которые синтезируют необходимые человеку вещества. Увеличение выхода продукта может быть достигнуто благодаря дальнейшей исследовательской работе по селекции специализированных популяций клеток и оптимизации условий культивирования. Большой интерес представляет также дальнейшее развитие методов биотрансформации метаболитов и иммобилизации культивируемых клеток.
Достаточно успешно развиваются с помощью технологий клеточной инженерии, культуры клеток и тканей ускорение и облегчение селекционного процесса, создание растений с новыми качествами, а также клональное микроразмножение растений, тесно связанное с проблемой их оздоровления от вирусных инфекций и криосохра-нение генофонда — технология, в настоящий момент приобретшая экологическую направленность.
Читайте также: Устьице тип ткани выполняемые функции
Дата добавления: 2016-06-02 ; просмотров: 2520 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Направления и возможности использования культуры изолированных тканей растений
Методы культивирования изолированных клеток, органов и тканей находят широкое применение в экспериментальной биологии и используются во многих биотехнологических процессах, в которые включены высшие растения.
Если биологической основой культуры изолированных тканей является тотипотентность растительных клеток, то обеспечение этого процесса складывается путем подбора питательных сред определенного состава, регуляторов роста и технических (физических) условий введения в культуру и собственно процесса культивирования (выбор экспланта, стерилизация, освещенность, температура и др.).
Основные направления использования
Культура клеток и тканей растений может использоваться в двух основных направлениях: 1) размножение или поддержание жизни у неизмененных по сравнению с донорами клеток, тканей, растений; 2) целенаправленное воздействие на изменение генетического статуса клеток и отбор в селективных условиях нужных вариантов.
Возможности культуры изолированных клеток и тканей растений весьма обширны:
1) получение вторичных метаболитов, продуцируемых отдельными клетками и тканями некоторых полезных растений (алкалоидов, глюкозидов, стероидов и т.д.), используемых для производства лекарств;
2) клеточная и тканевая селекция форм с полезными хозяйственными свойствами;
3) генетическое улучшение сельскохозяйственных растений;
4) ускоренное (микроразмножение) ценных и уникальных генотипов и поддержание жизнеспособности ослабленных клеток и тканей;
5) освобождение (оздоровление) посадочного материала от вирусной инфекции;
6) сохранение генофонда в условиях криоконсервации.
С использованием методов культивирования также решается большое число теоретических проблем, в том числе:
— особенности старения растительной клетки;
— процессы цитодифференцировки и морфогенеза;
— роль фитогормонов, углеводов, витаминов, минеральных веществ при каллусогенезе, органогенезе, морфогенезе;
— взаимоотношения клеток высших растений с клубеньковыми бактериями и микоризными грибами;
— механизмы устойчивости растений к неблагоприятным факторам среды: абиотическим (засолению, кислой среде, низким температурам и т.д.), биотическим (патогенам разного происхождения, вызывающим болезни растений);
— регуляция вторичного обмена;
— механизмы опухолеобразования;
— механизмы сомаклональной изменчивости;
— популяционные взаимоотношения в клеточной культуре.
Проблемы культивирования изолированных тканей
Успех в применении культуры клеток и тканей зависит от оптимизации физиологических процессов, обеспечивающих нормальное деление клеток, их дифференциацию и регенерацию из них взрослых растений, что определяется многими факторами, прежде всего используемыми гормонами. Наиболее сложной является регенерация растений из отдельных клеток. В первую очередь это касается злаковых растений, поэтому важнейшее значение имеет выяснение механизмов морфогенеза in vitro, регенерации и лежащих в их основе процессов.
Высокая способность к образованию растений-регенерантов при культивировании in vitro тканей и клеток растений является необходимым условием эффективного применения многих клеточных технологий.
В старых пересадочных культурах может протекать процесс, получивший название «привыкание». Он связан с усилением клеточной дедифференцировки, приобретением автономности по отношению к гормонам и превращением каллусных клеток в клетки химических опухолей. «Привыкание» тканей препятствует в большинстве случаев получению растений-регенерантов. Утрата регенерационной способности наблюдается также при длительном пассировании у гормонозависимых, т.е. нормальных, каллусных тканей.
Читайте также: Особенности распространения ультразвука в тканях
Новообразование растений в культивируемых каллусных культурах в значительной степени определяется генотипом и физиологическим статусом экспланта. В связи с этим способ отбора генотипов и эксплантов, эффективных к образованию морфогенетических структур в культуре in vitro, представляет собой актуальную задачу. Решение ее сопряжено с разработкой ряда физиологических проблем, одной из которых является изучение особенностей целого растения, обуславливающих поведение его органов и тканей в культуре in vitro.
Имеются данные, которые указывают на связь способности к регенерации растений в культуре первичных эксплантов с модификацией клеточного метаболизма, функционированием меристем и морфогенезом растения, которые, в свою очередь, определяются гормональным балансом.
Есть данные о положительном влиянии кустистости и об отрицательном влиянии скороспелости растений картофеля на число побегов в первичном каллусе клубневых дисков. Склонность к образованию многочисленных придаточных корней в природе находит отражение в высоком уровне ризогенеза в каллусной культуре. У гороха отмечен высокий коэффициент корреляции между азотфиксацией (нитрогеназная активность, интенсивность образования клубеньков, число клубеньков) и каллусо- и корнеобразованием.
Предполагается, что общая причина формирования клубеньков на корнях растений и образования каллуса в культуре in vitro заключается в способности генотипа к пролиферации клеток, определяемой балансом фитогормонов. Показана связь между полиэмбрионией семян in vivo и числом соматических эмбриоидов в культуре in vitro у ржи. Выявлено большое сходство при формировании эмбриоидов в семени пиона (Paeonia anomala L.) in vivo и в культуре in vitro (образование эмбриоидов из клеток эпидермиса, характер их развития, форма эмбриоидов и др.).
Отмечена отрицательная корреляция между частотой образования нередуцированных пыльцевых зерен и способностью к андрогенезу в культуре пыльников у картофеля. Возможно, это свидетельствует о сцеплении генов, контролирующих формирование эмбриоидов и первое деление при образовании реституционных ядер.
У раннеспелых сортов в культуре первичного каллуса, полученного из развивающихся зародышей, уровень множественной регенерации растений ниже, чем у позднеспелых. Одной из причин низкого уровня множественной регенерации растений в группе раннеспелых сортов может быть более быстрое развитие зародышей, что определяет более короткий период компетентности их клеток.
В работах, проведенных на злаках, показано влияние на процесс регенерации в культуре in vitro отдельных генов, вовлеченных в контроль гормонального баланса растений. Для пшеницы установлено влияние генов, обуславливающих короткостебельность, и гена, определяющего чувствительность к длине дня на рост каллуса, на соматический эмбриогенез и регенерацию растений. У ячменя выявлена сопряженность высокой регенерационной способности с числом рядов в колосе.
Известно, что древесные, и особенно хвойные, деревья характеризуются медленным ростом, трудно укореняются, содержат большое количество вторичных соединений (фенолы, терпены и другие вещества), которые в изолированных тканях активируются, окисляя фенолы растений, различные фенолазы. В свою очередь, продукты окисления фенолов обычно ингибируют деление и рост клеток, что ведет к гибели первичного экспланта или к уменьшению способности тканей древесных пород к регенерации адвентивных почек, которая с возрастом растения-донора постепенно исчезает полностью. Однако несмотря на все трудности, исследователи все чаще привлекают в качестве объектов исследований различные ткани и органы древесных растений.
В целом исследования влияния отдельных характеристик целого растения на поведение его частей в культуре in vitro малочисленны и фрагментарны.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом