
Гормоны, секретируемые железами внутренней секреции, связываются с транспортными белками плазмы или в некоторых случаях адсорбируются на клетках крови и доставляются к органам и тканям, влияя на их функцию и обмен веществ. Некоторые органы и ткани обладают очень высокой чувствительностью к гормонам, поэтому их называют органами-мишенямиилитканями–мишенями. Гормоны влияют буквально на все стороны обмена веществ, функции и структуры в организме.
Согласно современным представлениям, действие гормонов основано на стимуляции или угнетении каталитической функции определенных ферментов. Этот эффект достигается посредством активации или ингибирования уже имеющихся ферментов в клетках за счет ускорения их синтеза путём активации генов. Гормоны могут увеличивать или уменьшать проницаемость клеточных и субклеточных мембран для ферментов и других биологически активных веществ, благодаря чему облегчается или тормозится действие фермента.
Различают следующие типы механизма действия гормонов: мембранный, мембранно-внутриклеточный и внутриклеточный (цитозольный).
Мембранный механизм. Гормон связывается с клеточной мембраной и в месте связывания изменяет её проницаемость для глюкозы, аминокислот и некоторых ионов. В этом случае гормон выступает как эффектор транспортных средств мембраны. Такое действие оказывает инсулин, изменяя транспорт глюкозы. Но этот тип транспорта гормонов редко встречается в изолированном виде. Инсулин, например, обладает как мембранным, так и мембранно-внутриклеточным механизмом действия.
Мембранно-внутриклеточный механизм. По мембранно-внутриклеточному типу действуют гормоны, которые не проникают в клетку и поэтому влияют на обмен веществ через внутриклеточного химического посредника. К ним относят белково-пептидные гормоны (гормоны гипоталамуса, гипофиза, поджелудочной и паращитовидной желез, тиреокальцитонин щитовидной железы); производные аминокислот (гормоны мозгового слоя надпочечников – адреналин и норадреналин, щитовидной железы – тироксин, трийодтиронин).
Функции внутриклеточных химических посредников гормонов выполняют циклические нуклеотиды – циклический 3׳,5׳ – аденозинмонофосфат (цАМФ) и циклический 3׳,5׳ – гуанозинмонофосфат (цГМФ), ионы кальция.
Гормоны влияют на образование циклических нуклеотидов: цАМФ – через аденилатциклазу, цГМФ – через гуанилатциклазу.
Аденилатциклаза встроена в мембрану клетки и состоит из 3-х взаимосвязанных частей: рецепторной (R), представленной набором мембранных рецепторов, находящихся снаружи мембраны, сопрягающей (N), представленной особымN–белком, расположенным в липидном слое мембраны, и каталитической (C), являющейся ферментным белком, то есть собственно аденилатциклазой, которая превращает АТФ (аденозинтрифосфат) в цАМФ.
Аденилатциклаза работает по слудующей схеме. Как только гормон связывается с рецептором (R) и образуется комплекс гормон- рецептор, происходит образовагние комплексаN– белок – ГТФ (гуанозинтрифосфат), который активирует каталитическую (С) часть аденилатцеклазы. Активация аденилатциклазы приводит к образованию цАМФ внутри клетки на внутренней поверхности мембраны из АТФ.
Даже одна молекула гормона, связавшегося с рецептором, заставляет работать аденилатцеклазу. При этом на одну молекулу связавшегося гормона образуется 10-100 молекул цАМФ внутри клетки. В активном состоянии аденилатциклаза находится до тех пор, пока существует комплекс гормон – рецептор. Аналогичным образом работает и гуанилатциклаза.
В цитоплазме клетки находятся неактивные протеинкиназы. Циклические нуклеотиды- цАМФ ицГМФ- активируют пртеинкиназы. Существуют цАМФ- зависимые и цГМФ – зависимые протеинкиназы, которые активируются своим циклическим нуклеотидом. В зависимости от мембранного рецептора, связывающего определенный гормон, включается или аденилатцеклаза, или гуанилатцеклазаи соответственно происходит образование или цАМФ, или цГМФ.
Через цАМФ действует большинство гормонов, а через цГМФ- только окситоцин, тиреокальцитонин, инсулин и адреналин.
При помощи активированных протеинкиназ осуществляется два вида регуляции активности ферментов: активация уже имеющихся ферментов путем ковалентной модификации, то есть фосфолированием; изменение количества ферментного белка за счет изменения скорости его биосинтеза.
Влияние циклических нуклеотидов на биохимические процессы прекращается под влиянием специального фермента- фосфодиэстеразы, разрушающей цАМФ и цГМФ. Другой фермент – фосфопротеидфосфаза – разрушает результат действия протеинкиназы, то есть отщепляет фосфорную кислоту от ферментных белков, в результате чего они становятся неактивными.
Внутри клетки ионов кальция содержится очень мало, вне клетки их больше. Они пступают из внеклеточной среды по кальциевым каналам в мембране. В клетке кальций взаимодействует с кальцийсвязывающим белком калмодулином (КМ). Этот комплекс изменяет активность ферментов, что ведет к изменению физиологический функций клеток. Через ионы кальция действуют гормоны окситоцин, инсулин, простагландин F2α. Таким образом, чувствительность тканей и органов к гормонам зависит от мембранных рецепторов, а специфическое регуляторное влияние их определяется внутриклеточным посредником.
Читайте также: Dikline kd31 ткань elain 28 фуксия
Внутриклеточный (цитозольный) механизм действия. Он характерен для стероидных гармонов (кортикостероидов, половых гормонов – андрогенов, эстрогенов и гестагенов). Стероидгные гормоны взаимодействуют с рецепторами, находящимися в цитоплазме. Образовавшийся гормнон-рецепторный комплекс переносится в ядро и действует непосредственно на геном, стимулируя или угнетая его активность, т.е. действует на синтез ДНК, изменяя скорость транскрипции и количество инфармационной (матричной) РНК (мРНК). Увеличение или уменьшение количества мРНК влияет на синтез белка в процессе трансляции, что приводит к изменению функциональной активности клетки.
Механизмы действия гормонов на клетки-мишени.
В зависимости от строения гормона существуют два типа взаимодействия. Если молекула гормона липофильна, (например, стероидные гормоны), то она может проникать через липидный слой наружной мембраны клеток-мишеней. Если молекула имеет большие размеры или является полярной, то ее проникновение внутрь клетки невозможно. Поэтому для липофильных гормонов рецепторы находятся внутри клеток-мишеней, а для гидрофильных — рецепторы находятся в наружной мембране.
Для получения клеточного ответа на гормональный сигнал в случае гидрофильных молекул действует внутриклеточный механизм передачи сигнала. Это происходит с участием веществ, которых называют «ВТОРЫМИ ПОСРЕДНИКАМИ». Молекулы гормонов очень разнообразны по форме, а «вторые посредники» — нет.
Надёжность передачи сигнала обеспечивает очень высокое сродство гормона к своему белку-рецептору.
Что такое посредники, которые участвуют во внутриклеточной передаче гуморальных сигналов? Это циклические нуклеотиды (цАМФ и цГМФ), инозитолтрифосфат, кальций-связывающий белок — кальмодулин, ионы кальция, ферменты, участвующие в синтезе циклических нуклеотидов, а также протеинкиназы — ферменты фосфорилирования белков. Все эти вещества участвуют в регуляции активности отдельных ферментных систем в клетках-мишенях.
Разберем более подробно механизмы действия гормонов и внутриклеточных посредников. Существует два главных способа передачи сигнала в клетки-мишени от сигнальных молекул с мембранным механизмом действия:
1. АДЕНИЛАТЦИКЛАЗНАЯ (ИЛИ ГУАНИЛАТЦИКЛАЗНАЯ) СИСТЕМЫ
2. ФОСФОИНОЗИТИДНЫЙ МЕХАНИЗМ
Аденилатциклазная система.
Основные компоненты: мембранный белок-рецептор, G-белок, фермент аденилатциклаза, гуанозинтрифосфат, протеинкиназы.
Кроме того, для нормального функционирования аденилатциклазной системы, требуется АТФ.
Схема аденилатциклазной системы представлена на рисунке:
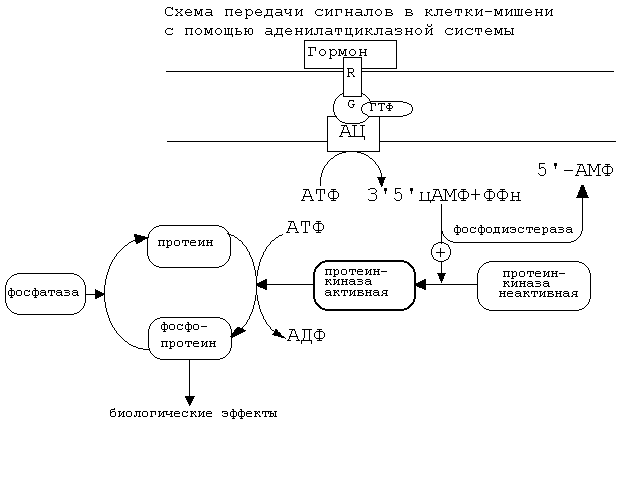
Как видно из рисунка, белок-рецептор, G-белок, рядом с которым располагаются ГТФ и фермент (аденилатциклаза) встроены в мембрану клетки.
До момента действия гормона эти компоненты находятся в диссоциированнном состоянии, а после образования комплекса сигнальной молекулы с белком-рецептором происходят изменения конформации G-белка. В результате одна из субъединиц G-белка приобретает способность связываться с ГТФ.
Комплекс “G-белок-ГТФ” активирует аденилатциклазу. Аденилатциклаза начинает активно превращать молекулы АТФ в ц-АМФ.
ц-АМФ обладает способностью активировать особые ферменты — протеинкиназы, которые катализируют реакции фосфорилирования различных белков с участием АТФ. При этом в состав белковых молекул включаются остатки фосфорной кислоты. Главным результатом этого процесса фосфорилирования является изменение активности фосфорилированного белка. В различных типах клеток фосфорилированию в результате активации аденилат-циклазной системы подвергаются белки с разной функциональной активностью. Например, это могут быть ферменты, ядерные белки, мембранные белки. В результате реакции фосфорилирования белки могут становятся функционально активными или неактивными.
Такие процессы будут приводить к изменениям скорости биохимических процессов в клетке-мишени.
Активация аденилатциклазной систтемы длится очень короткое время, потому что G-белок после связывания с аденилатциклазой начинает проявлять ГТФ-азную активность. После гидролиза ГТФ G-белок восстанавливает свою конформацию и перестает активировать аденилатциклазу. В результате прекращается реакция образования цАМФ.
Кроме участников аденилатциклазной системы в некоторых клетках-мишенях имеются белки-рецепторы, связанные с G-белками, которые приводят к торможению аденилатциклазы. При этом комплекс “GTP-G-белок” ингибирует аденилатциклазу.
Когда останавливается образование цАМФ, реакции фосфорилирования в клетке прекращаются не сразу: пока продолжают существовать молекулы цАМФ — будет продолжаться и процесс активации протеинкиназ. Для того, чтобы прекратить действие цАМФ, в клетках существует специальный фермент — фосфодиэстераза, который катализирует реакцию гидролиза 3′,5′-цикло-АМФ до АМФ.
Читайте также: Как правильно сшивать лоскутки ткани
Некоторые вещества, обладающие ингибирующим действием на фосфодиэстеразу, (например, алкалоиды кофеин, теофиллин), способствуют сохранению и увеличению концентрации цикло-АМФ в клетке. Под действием этих веществ в организме продолжительность активации аденилатциклазной системы становится больше, то есть усиливается действие гормона.
Кроме аденилат-циклазной или гуанилатциклазной систем существует также механизм передачи информации внутри клетки-мишени с участием ионов кальция и инозитолтрифосфата.
Инозитолтрифосфат -это вещество, которое является производным сложного липида — инозитфосфатида. Оно образуется в результате действия специального фермента — фосфолипазы “С”, который активируется в результате конформационных изменений внутриклеточного домена мембранного белка-рецептора.
Этот фермент гидролизует фосфоэфирную связь в молекуле фосфатидил-инозитол-4,5-бисфосфата и в результате образуются диацилглицерин и инозитолтрифосфат.

Известно, что образование диацилглицерина и инозитолтрифосфата приводит к увеличению концентрации ионизированного кальция внутри клетки. Это приводит к активации многих кальций-зависимых белков внутри клетки, в том числе активируются различные протеинкиназы. И здесь, как и при активации аденилатциклазной системы, одной из стадий передачи сигнала внутри клетки является фосфорилирование белков, которое в приводит к физиологическому ответу клетки на действие гормона.
В работе фосфоинозитидного механизма передачи сигналов в клетке-мишени принимает участие специальный кальций-связывающий белок — кальмодулин. Это низкомолекулярный белок (17 кДа), на 30% состоящий из отрицательно заряженных аминокислот (Глу, Асп) и поэтому способный активно связывать Са +2 . Одна молекула кальмодулина имеет 4 кальций-связывающих участка. После взаимодействия с Са +2 происходят конформационные изменения молекулы кальмодулина и комплекс “Са +2 -кальмодулин” становится способным регулировать активность (аллостерически угнетать или активировать) многие ферменты — аденилатциклазу, фосфодиэстеразу, Са +2 ,Мg +2 -АТФазу и различные протеинкиназы.
В разных клетках при воздействии комплекса “Са +2 -кальмодулин” на изоферменты одного и того же фермента (например, на аденилатциклазу разного типа) в одних случаях наблюдается активация, а в других — ингибирование реакции образования цАМФ. Такие различные эффекты происходят потому, что аллостерические центры изоферментов могут включать в себя различные радикалы аминокислот и их реакция на действие комплекса Са +2 -кальмодулин будет отличаться.
Таким образом, в роли «вторых посредников» для передачи сигналов от гормонов в клетках-мишенях могут быть:
— Циклические нуклеотиды (ц-АМФ и ц-ГМФ);
Механизмы передачи информации от гормонов внутри клеток-мишеней с помощью перечисленных посредников имеют общие черты:
1. одним из этапов передачи сигнала является фосфорилирование белков
2. прекращение активации происходит в результате специальных механизмов, инициируемых самими участниками процессов, — существуют механизмы отрицательной обратной связи.
Гормоны являются основными гуморальными регуляторами физиологических функций организма, и в настоящее время хорошо известны их свойства, процессы биосинтеза и механизмы действия.
Признаки, по которым гормоны отличаются от других сигнальных молекул:
1. Синтез гормонов происходит в особых клетках эндокринной системы. При этом синтез гормонов является основной функцией эндокринных клеток.
2. Гормоны секретируются в кровь, чаще в венозную, иногда в лимфу. Другие сигнальные молекулы могут достигать клеток-мишеней без секреции в циркулирующие жидкости.
3. Телекринный эффект (или дистантное действие) — гормоны действуют на клетки-мишени на больщом расстоянии от места синтеза.
Гормоны являются высокоспецифичными веществами по отношению к клеткам-мишеням и обладают очень высокой биологической активностью.
1.2. Механизмы действия гормонов на клетки — мишени
Конечными эффектами действия гормонов на клеточном уровне могут быть изменения метаболизма, проницаемости мембраны для различных ве ществ (ионов, глюкозы и др.), процессов роста, дифференцировки и деления клеток, сократительной или секреторной активности и др. Реализация этих эффектов начинается со связывания гормона со специфическими клеточны ми белками-рецепт орами : мембранными или внутриклеточными (цито плазматическими и ядерными). Эффект действия гормонов через мембранные рецепторы проявляется относительно быстро (в течение нескольких минут), а через внутриклеточные рецепторы — медленно (от получаса и более).
Действие через м ем бранны е рецепторы типично для белково-пептид ных гормонов и производных аминокислот. Эти гормоны (за исключением тироидных), являются гидрофильными и не могут проникать сквозь били пидный слой плазмалеммы. Поэтому, гормональный сигнал передается в клетку по относительно длинной цепи, которая в общем случае выглядит так: гормон —> мембранный рецептор —> мембранный фермент —> вторичный посредник —> протеинкиназа —> внутриклеточные функциональные белки —> физиологический эффект.
Читайте также: Как сделать ткань крэш в домашних условиях
Соответственно действие гормона через мембранные рецепторы реа лизуется в несколько этапов:
1 ) взаимодействие гормона с мембранным рецептором приводит к изменению конформации рецептора и его активированию;
2) рецептор активирует (реже — ингибирует) связанный с ним мем бранный фермент ;
3) фермент изменяет концентрацию в цитоплазме того или иного низкомолекулярного вещества — вторичного посредника’,
4) вторичный посредник активирует определенную цитоплазматичес кую протеинкиназу — фермент, катализирующий фосфорилирование и из менение функциональных свойств белков;
5) протеинкиназа изменяет активность внутриклеточных функцио нальных белков, регулирующих внутриклеточные процессы (ферментов, ионных каналов, сократительных белков и т.п.), в результате чего реализу ется тот или иной конечный эффект действия гормона, например, ускоре ние синтеза или распада гликогена, запуск мышечного сокращения и др.
В настоящее время известны четыре типа ферментов, связанных с мембранными рецепторами гормонов, и пять основных вторичных посредников (рис. 1, табл. 1).
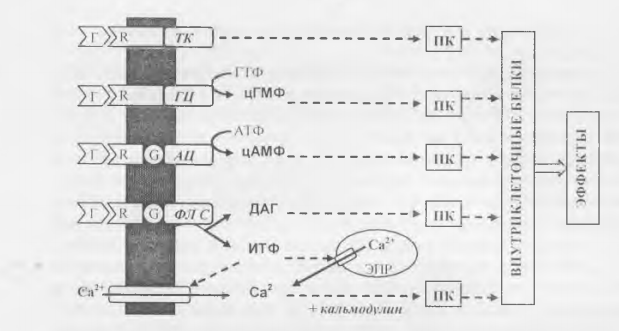
Рис. 1. Основные системы трансмембранной передачи гормонального сигнала.
Обозначения: Г — гормоны; R — мембранные рецепторы; G — G-белки; Ж — тирозин-
киназа; Г Ц — гуанилатциклаза; А Ц
аденилатциклаза; Ф.П С — фосфолипаза С; фл — мембранные фосфолипиды; ИТФ — инозитолтрифосфат, Д АТ — диацилглицерол; ЭПР — эндоплазматический ретикулум; ПК — различные протеинкиназы.
Мембранные ферменты и вторичные посредники, опосредующие действие гормонов через мембранные рецепторы
Основные активирующие гормоны
инсулин, гормон роста, пролактин
предсердный натрийуретический гормон
многие гормоны, например, адреналин через 3-адренорецепторы
многие гормоны, например, адреналин через а г адренорецепторы
В зависимости от того, как осуществляется связь между рецептороми мембранным ферментом различают два типа рецепторов: 1) каталитические рецепторы; 2)рецепторы, сопряженные с G-белками.
Каталитические рецепторы: рецептор и фермент связаны непосредственно (могут представлять собой одну молекулу с двумя функциональными участками). Мембранными ферментами у этих рецепторов могут быть:
• тирозинкиназа (разновидность протеинкиназ); действие гормонов через тирозинкиназные рецепторы не требует обязательного наличия вторичных посредников;
• гуанилатциклаза — катализирует образование вторичного посредника циклической ГМФ (цГМФ) из ГТФ.
Рецепторы, сопряженные с G-белками: сигнал от молекулы рецептора передается сначала особому мембранному G-белку1, который далее активирует или ингибирует определенный мембранный фермент, которым может быть:
• аденилатциклаза — катализирует образование вторичного посредника циклической АМФ (цАМФ) из АТФ;
• фосфолипаза С — катализирует образование из мембранных фосфолипидов двух вторичных посредников: инозитолтрифосфата (ИТФ) и диацилглицерола (ДАГ). ДАГ стимулирует протеинкиназу, а также является предшественником простагландинов и подобных им биологически активных веществ. Основной эффект ИТФ заключается в повышении содержания в цитоплазме еще одного вторичного посредника — ионов Са 2+ , которые поступают в цитозоль через ионные каналы плазмалеммы (из внеклеточной среды) или внутриклеточных депо Са2+(эндоплазматического ретикулума и митохондрий). Свое физиологическое действие ионы Са2+ осуществляют, как правило, в соединении с белком кальмодулином.
Действие через внутриклеточные рецепторы типично для стероидных и тироидных гормонов, которые благодаря своей жирорастворимости способны проникать через клеточные мембраны внутрь клетки и ее ядра (рис. 2).
Взаимодействуя с ядерными рецепторами, указанные гормоны влияют на процессы клеточного деления и реализации генетической информации (экспрессии генов), в частности, регулируют скорость биосинтеза функциональных клеточных белков — ферментов, рецепторов, пептидных гормонов и др.
В результате действия гормонов на цитоплазматические рецепторы изменяется активность клеточных органелл, например, интенсивность биологического окисления в митохондриях или синтеза белка в рибосомах.
В комплексе с цитоплазматическими рецепторами гормоны могут прони кать в ядро, действуя так же, как через ядерные рецепторы.
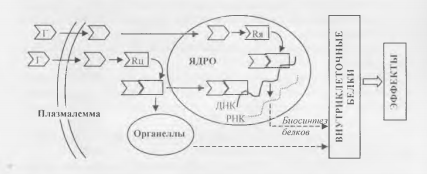
Рис.2. Механизмы внутриклеточного действия гормонов.
Обозначения: Г — гормоны; Rh — ядерные рецепторы; Rif — цитоплазматические рецепторы.