
Общие пути обмена аминокислот. Пути обезвреживания аммиака в организме
Реферат выполнила студентка 12 группы II курса пед/ф-та Пасько С.П..
Саратовский Государственный Медицинский Университет
Промежуточный обмен аминокислот в тканях.
Промежуточный метаболизм аминокислот белковых молекул, как и других питательных веществ в организме, включает катаболические (распад до конечных продуктов) и анаболические (биосинтез аминокислот) процессы, а также ряд других специфических превращений, сопровождающихся образованием биологически активных веществ. Условно промежуточный метаболизм аминокислот можно разделить на общие пути обмена и индивидуальные превращения отдельных аминокислот.
Общие пути обмена аминокислот.
Общие пути превращения аминокислот включают реакции дезаминирования, трансаминирования, декарбоксилирования, биосинтеза и рацемизации. Реакции рацемизации характерны только для микроорганизмов, физиологическая роль которой заключается в синтезе D-изомеров аминокислот для построения клеточной оболочки.
Дезаминирование ( отщепление аминогруппы) – существует четыре типа реакций, катализируемых своими ферментами:
Восстановительное дезаминорование ( +2H + )
Гидролитическое дезаминированиие (+H 2 О)
Окислительное дезаминирование (+1/2 О 2 )
Во всех случаях NH 2 — группа аминокислоты высвобождается в виде аммиака. Помимо аммиака продуктами дезаминирования являются жирные кислоты, окикислоты и кетокислоты. Для животных тканей, растений и большинства микроорганизмов преобладающим типом реакций является окислительное дезаминирование аминокислот, за исключением гистидина, который подвергается внутримолекулярному дезаминированию.
Кроме перечисленных четырех типов реакций и катализирующих их ферментов в животных тканях и печени человека открыты также три специфических фермента (серин- и треониндегидратазы и цистатионин-γ- лиаза), катализирующих неокислительное дезаминирование серина, треонина и цистеина. Они требуют присутствия пиридоксаль-фосфата в качестве кофермента. Конечными продуктами реакции являются пируват и α- кетобутират, аммиак и сероводород.
Трансаминирование – реакции межмолекулярного переноса аминогруппы (NH 2 ) от аминокислоты на α-кетокислоту без промежуточного образования аммиака (глутамат+ пируват =
α-кетоглутарат + аланин). Впервые эти реакции были открыты в 1937г. А.Е. Браунштейном и М.Г. Крицман. Реакции трансаминирования являются обратимыми и универсальными для всех живых организмов, они протекают при участии специфических ферментов – аминотрансфераз (трансамниназ). Теоретически реакции возможны между любой амино- и кетокислотой, но наиболее интенсивно они протекают, если один из партнеров представлен дикарбоновой амино- или кетокислотой. В переносе амниогруппы активное участие принимает кофермет трансминаз – пиридоксальфосфат (производное витамина В 6 ). Для реакций трансаминирования характерен общий механизм. Ферменты реакции катализируют перенос аминогруппы не на α -кетокислоту, а на кофермент; образовавшееся промежуточное соединение (шиффово основание) подвергается внутримолекулярным превращениям, приводящим к освобождению α-кетокислоты и пиридоксамнофосфата. Последний на втолрой стадии реагирует с любой другой α-кетокислотой, что через те же стадии приводит к синтезу новой аминокислоты и пиридоксальфосфата.
Декарбоксилирование — отщепление карбоксильной группы в виде СО 2 , образующиеся продукты реакции называются биогенными аминами, они оказывают сильное фармакологическое действие на множество функций. Эти реакции являются необратимыми, они катализируютя специфическими ферментами – декарбоксилазами аминокмлот- которые в качестве кофермента содержат пиридоксальфосфат ( кроме гистидиндекарбоксилазы и аденозилдекарбоксилазы – содержат остаток пировиноградной кислоты в качестве кофермента). В живых организмах открыты четыре типа декарбоксилирования аминокислот.
α-декарбоксилирование – характерно для тканей животных: от аминокислот отщепляется соседняя от α-углеродного атома карбоксильная группа.
ω-декарбоксилирование- свойственно микроорганизмам
декарбоксилирование, связанное с реакцией трансаминирования. Образуется альдегид и новая аминокислота, соответствующая исходной кетокислоте.
Декарбоксилирование, связанное с реакцией конденсацией двух молекул:
Обезвреживание аммиака в организме.
В организме человека подвергается распаду около 70г аминокислот в сутки: при этом освобождается большое количество аммиака, являющегося высокотоксичным соединением. Поэтому крнцентрация аммиака должна сохраняться на низком уровне (в норме уровень его не превышает 60 мкмоль/л). Концентрация аммиака 3 ммоль/л является летальной.
Одним из путей связывания и обезвреживания аммиака в мозге, сетчатке, почках и мышцах, является биосинтез глутамина( и, возможно, аспарагина). Поскольку глутамин и аспарагин с мочой выделяются в небольших количествах, было высказано предположение, что они выполняют скорее транспортную функцию переноса аммиака в нетоксичной форме.
Часть аммиака легко связывается с α-кетоглутаровой кислотой благодаря обратимости глутаматдегидрогеназной реакции; при синтезе глутамина связывается ещё 1 молекула, т.о. нейтрализуются две молекулы аммиака:
Орнитиновый цикл мочевинообразования.
Основным механизмом обезвреживания аммиака в организме является биосинтез мочевины (в основном,в печени).Она выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного, обмена. На долю мочевины приходится до 80-85% всего азота мочи. Реакции синтеза мочевины, представлены в виде цикла, получившего название орнитинового цикла мочевинообразования Кребса.
На первом этапе синтезируется макроэргическое соединение карбамоилфосфат – это метаболически активная форма аммиака, используемая в качестве исходного продукта для синтеза ряда других азотистых соединений.
На втором этапе цикла мочевинообразования происходит конденсация карбамоилфосфата и орнитина с образованием
Читайте также: Навес из ткани для окон
цитруллина; реакцию катализирует орнитинкарбамоилтрансфераза:
На следующей стадии цитруллин превращается в аргинин в результате двух последовательно протекающих реакций. Первая из них, энергозависимая, сводится к конденсации цитруллина и аспаргиновой кислоты с образованием аргининосукцината ( эту реакцию катализирует аргининосукцинат-синтетаза). Аргининсукцинат распадается во второй реакции на аргинин и фумарат поддействием аргининосукцинат-лиазы.
На последнем этапе аргинин расщепляется на мочевину и орнитин под действием аргиназы. Суммарная реакция синтеза мочевины без учёта промежуточных продуктов:
Это энергетически выгодная реакция, поэтому процесс всегда протекает в направлении синтеза мочевины.
В состоянии азотистого равновесия организм человека потребляет и соответственно выделяет примерно 15 г азота в сутки; из экскретируемого с мочой количества азота на долю мочевины приходится около 85% , креатинина-около 5%, аммонийных солей – 3%, мочевой кислоты-1% и на другие формы-около 6%.
Типы азотистого обмена. А м м о н и о т е л и ч е с к и й т и п , при котором главным конечным продуктом азотистого обмена является аммиак, свойствен рыбам. У р е о т е л и ч е с к и й т и п обмена — основным конечным продуктом обмена белков является мочевина, характерен для человека и животных. У р и к о т е л и ч е с к и й т и п — главным конечным продуктом обмена является мочевая кислота, характерен для птиц и рептилий.
Пути обезвреживания аммиака в тканях животных
Аммиак токсичен, и накопление его в организме может привести к смерти. Существуют следующие пути обезвреживания аммиака:
1. Синтез аммонийных солей.
2. Синтез амидов дикарбоновых аминокислот.
Синтез аммонийных солей происходит ограниченно в почках, это как дополнительное защитное приспособление организма при ацидозах. Аммиак и кетокислоты частично используются для ресинтеза аминокислот и для синтеза других азотистых веществ. Кроме того, в тканях почек аммиак участвует в процессе обезвреживания оргинических и неорганических кислот, образуя с ними нейтральные и кислые соли:
Этим путем организм защищается от потери с мочой при выведениикислот знпачительного количества катионов (Na, K, отчасти Са, Mg), что могло бы привести к резкому снижению щелочного резерва крови. Количество аммонийных солей, выводимых с мочой, заметно повышается при ацидозе, так как аммиак используется для нейтрализации кислоты. Одним из путей связывания и обезвреживания аммиака является использование его для образования амидной связи глутамина и аспарагина. При этом из глутаминовой кислоты под действием фермента глутаминсинтетазы синтезируется глутамин, из аспарагиновой кислоты при участии аспарагинсинтетазы – аспарагин:
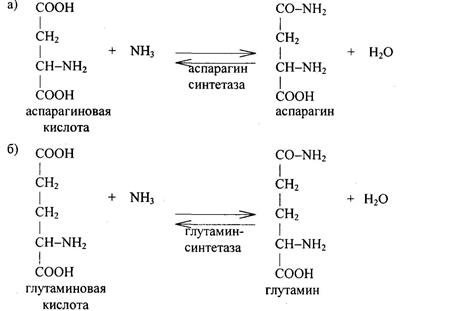
Этим путем происходит устранение аммиака во многих органах (мозг, сетчатка, почки, печень, мышцы). Амиды глутаминовой и аспарагиновой кислот могут образоваться и тогда, когда эти аминокислоты находятся в структуре белка, то есть акцептором аммиака может быть не только свободная аминокислота, но и белки, в состав которых они входят. Аспарагин и глутамин доставляются в печень и используются в синтезе мочевины. Аммиак переносится в печень и с помощью аланина (глюкозо-аланиновый цикл). Этот цикл обеспечивает перенос аминогрупп из скелетных мышц в печень, где они превращаются в мочевину, а работающие мышцы получают глюкозу. В печени глюкоза синтезируется из углеродного скелета аланина. В работающей мышце из α-кетоглутаровой кислоты образуется глутаминовая кислота, которая затем передает аминную группу — NH2 пировиноградной кислоте, в результате синтезируется аланин – нейтральная аминокислота. Схематически указанный цикл выглядит следующим образом:
Глутаминовая кислота + пировиноградная кислота ↔
↔ α-кетоглутаровая кислота + аланин
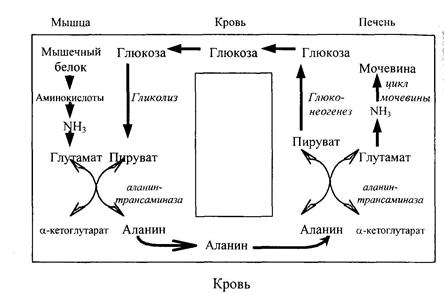
Рис. 10.1. Глюкозо-аланиновый цикл.
Этот цикл выполняет две функции: 1) переносит аминогруппы из скелетных мышц в печень, где они превращаются в мочевину;
2) обеспечивает работающие мышцы глюкозой, поступающей с кровью из печени, где для ее образования используется углеродный скелет аланина.
Образование мочевины – основной путь обезвреживания аммиака. Этот процесс изучали в лаборатории И.П.Павлова. Показано, что мочевина синтезируется в печени из аммиака, CO2 и воды.
Мочевина выводится с мочой в качестве главного конечного продукта белкового, соответственно аминокислотного обмена. На долю мочевины приходится до 80-85% всего азота мочи. Главным местом синтеза мочевины в организме является печень. Сейчас доказано, что синтез мочевины происходит в несколько этапов.
1 стадия – образование карбамоилфосфата происходит в митохондриях под действием фермента карбомоилфосфат-синтетазы:
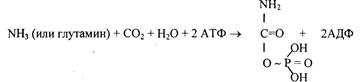
На следующей стадии с участием орнитина синтезируется цитруллин:
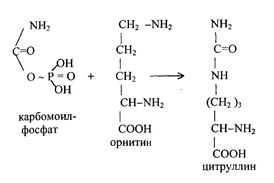
Цитруллин переходит из митохондрий в цитозоль клеток печени. После этого в цикл вводится вторая аминогруппа в форме аспарагиновой кислоты. Происходит конденсация молекул цитруллина и аспарагиновой кислоты с образованием аргинин-янтарной кислоты.
Читайте также: Акварель пэчворк как резать ткань
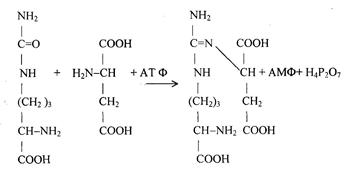
Цитруллин аспарагиновая аргинин-янтарная
Аргинин-янтарная кислота расщепляется на аргинин и фумаровую кислоты.
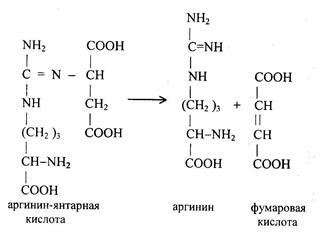
Под действием аргиназы аргинин гидролизуется, образуется мочевина и орнитин. В дальнейшем орнитин поступает в митохондрии и может включиться в новый цикл обезвреживания аммиака, а мочевина выделяется с мочой.
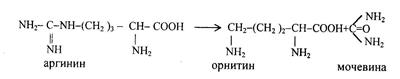
Таким образом, в синтезе одной молекулы мочевины нейтрализуется две молекулы NH3 и CO2 (HCO3), что также имеет значение в поддержании рН. Для синтеза одной молекулы мочевины расходуется 3 молекулы АТФ, в том числе две при синтезе карбомоилфосфата, одна для образования аргинин-янтарной кислоты; фумаровая кислота может превращаться в яблочную и щавелевоуксусную кислоты (цикл Кребса), а последняя в результате трансаминирования или восстановительного аминирования может превратиться в аспарагиновую кислоту. Некоторая часть азота аминокислот выделяется из организма в виде креатинина, который образуется из креатина и креатинфосфата.
Из всего азота мочи на долю мочевины приходится до 80-90%, аммонийных солей – 6 %. При избыточном кормлении белком доля азота мочевины возрастает, а при недостаточном белковом кормлении снижается до 60 %.
У птиц и рептилий – нейтрализация аммиака происходит путем образования мочевой кислоты. Птичий помет на птицефабриках — это источник азотсодержащего удобрения (мочевая кислота).
Образование и обезвреживание аммиака
Тема: «ОБРАЗОВАНИЕ АММИАКА В ОРГАНИЗМЕ И ПУТИ ЕГО ОБЕЗВРЕЖИВАНИЯ»
1. Источники и пути образования аммиака в организме. Возможные механизмы токсического действия аммиака.
2. Обезвреживание аммиака в тканях. Транспортные формы аммиака в крови (глутамин, аспарагин, аланин), их роль в организме.
3. Выведение аммиака почками: ферменты, участвующие в этом процессе, механизм транспорта, роль в организме.
4. Биосинтез мочевины в печени: локализация в клетке, ферменты, промежуточные продукты, энергетический баланс, регуляция, роль в организме.
5. Взаимосвязь цикла мочевинообразования и цикла трикарбоновых кислот.
6. Генетические дефекты ферментов цикла мочевинообразования: основные симптомы заболеваний, биохимическая диагностика, особенности диеты.
7. Клинико-диагностическое значение определения содержания мочевины в крови и моче.
Источники и пути образования аммиака.
24.1.1. Аммиак (NН 3 ) – продукт обмена большинства соединений, содержащих амино- и амидогруппы. Главным путём образования аммиака служит окислительное дезаминирование.
Аммиак – очень токсичное вещество, особенно для нервной системы. При физиологических значениях рН молекула NН 3 легко превращается в ион аммония NН 4 + , который не способен проникать через биологические мембраны и задерживается в клетке. Накопление NН 4 + вызывает торможение заключительных этапов цикла трикарбоновых кислот и снижение продукции АТФ. Поэтому в организме существует ряд механизмов связывания (обезвреживания) аммиака (см. рисунок 24.1).
Рисунок 24.1. Образование аммиака в организме и его обезвреживание.
Образование транспортных форм аммиака в тканях.
24.2.1 Аммиак, образующийся в тканях, сначала превращается в нетоксичное соединение и в таком виде переносится кровью к печени или почкам. Такими транспортными формами являются аминокислоты глутамин, аспарагин и аланин.
24.2.2. Образование глутамина и аспарагина из глутамата и аспартата соответственно происходит во многих тканях, включая головной мозг:
Глутамин — нейтральное нетоксичное соединение, способное легко проходить через клеточные мембраны. В виде этой аминокислоты аммиак транспортируется в крови. В крови здоровых людей содержание глутамина существенно превышает содержание других аминокислот. Глутамин, помимо участия в синтезе белка, служит источником азота в биосинтезе гистидина, глюкозамина, пуриновых и пиримидиновых нуклеотидов. С кровью глутамин поступает в печень и почки. Здесь он под действием фермента глутаминазы превращается в глутамат и аммиак. При участии аспарагиназы также происходит образование аммиака из аспарагина.
24.2.3. Аланин является транспортной формой аммиака, которая образуется преимущественно в мышцах. При интенсивной физической нагрузке источниками аммиака служат реакции дезаминирования аминокислот и аденозинмонофосфата (АМФ). Сначала аммиак превращается в аминогруппу глутамата в реакции восстановительного аминирования , катализируемой глутаматдегидрогеназой(см. параграф 18.6.2):
Образовавшийся глутамат переносит затем свою α-аминогруппу на пируват, всегда имеющийся в достаточном количестве, поскольку это продукт протекающего в мышцах гликолиза. Реакция катализируется аланинаминотрансферазой.
Глутамат + Пируват α-Кетоглутарат + Аланин
Аланин (нейтральная аминокислота, не несущая суммарного заряда при значениях рН, близких к 7) выходит из клеток и доставляется кровью к печени. Здесь он под действием аланинаминотрансферазы передаёт свою аминогруппу α-кетоглутарату, в результате чего образуется глутамат.
α-Кетоглутарат + Аланин Глутамат + Пируват
Далее в реакции, катализируемой глутаматдегидрогеназой, глутамат дезаминируется с образованием α-кетоглутарата и аммиака, который в печени превращается в мочевину.
Экскреция аммиака с мочой.
24.3.1. Клетки почек поглощают из циркулирующей крови глутамин. Фермент глутаминаза в почках катализирует гидролиз глутамина с образованием глутамата и аммиака.
Образующийся глутамат может в дальнейшем подвергаться дезаминированию при участии глутаматдегидрогеназы. Таким образом, из одной молекулы глутамина всего может образоваться две молекулы аммиака.
Читайте также: Кошка сплюшка из ткани
Аспарагин аналогичным образом подвергается гидролизу при участии аспарагиназы, присутствующей в клетках канальцев почек.
24.3.2. Аммиак секретируется в просвет почечных канальцев и выводится в виде солей минеральных и органических кислот, например:
NН 3 + НСl = NН 4 Сl
хлорид аммония
Образование солей аммония в почечных канальцах является важным механизмом регуляции кислотно-основного состояния организма. Оно резко возрастает при метаболическом ацидозе — накоплении в организме кислот и снижается при потере кислот организмом (алкалозе).
Биосинтез мочевины.
24.4.1. Биосинтез мочевины – основной путь обезвреживания аммиака. Мочевина синтезируется в орнитиновом цикле, протекающем в клетках печени. Эту последовательность реакций открыли Х.Кребс и К.Хензелейт в 1932 г. Согласно современным представлениям, цикл мочевины включает последовательность пяти реакций.
Две начальные реакции биосинтеза мочевины происходят в митохондриях клеток печени.
Последующие реакции протекают в цитоплазме клеток печени.
Общая схема орнитинового цикла представлена на рисунке 24.2:
Рисунок 24.2. Схема орнитинового цикла и его связь с превращениями фумаровой и аспарагиновой кислот.
Цифрами обозначены ферменты, катализирующие реакции орнитинового цикла: 1 – карбамоилфосфатсинтетаза; 2 – орнитин-карбамоилтрансфераза; 3 – аргининосукцинатсинтетаза; 4 – аргининосукцинатлиаза; 5 – аргиназа.
24.4.2. Орнитиновый цикл находится в тесной взаимосвязи с циклом трикарбоновых кислот:
- пусковые реакции цикла мочевины, как и реакции ЦТК, протекают в митохондриальном матриксе;
- поступление СО 2 и АТФ, необходимых для образования мочевины, обеспечивается работой ЦТК;
- в цикле мочевины образуется фумарат, который является одним из субстратов ЦТК. Фумарат гидратируется в малат, который в свою очередь окисляется в оксалоацетат. Оксалоацетат может подвергаться трансаминированию в аспартат; эта аминокислота участвует в образовании аргининосукцината.
24.4.3. Регуляция активности ферментов цикла осуществляется главным образом на уровне карбамоилфосфатсинтетазы, которая малоактивна в отсутствие своего аллостерического активатора — N-ацетил-глутамата. Концентрация последнего зависит от концентрации его предшественников (ацетил-КоА и глутамата), а также аргинина, который является аллостерическим активатором N-ацетилглутаматсинтазы:
Ацетил-КоА + Глутамат N-ацетилглутамат + КоА-SH
Концентрация ферментов орнитинового цикла зависит от содержания белка в пищевом рационе. При переходе на диету, богатую белком, в печени повышается синтез ферментов орнитинового цикла. При возвращении к сбалансированному рациону концентрация ферментов снижается. В условиях голодания, когда усиливается распад тканевых белков и использование аминокислот как энергетических субстратов, возрастает продукция аммиака, концентрация ферментов орнитинового цикла увеличивается.
24.4.4. Нарушения орнитинового цикла. Известны метаболические нарушения, обусловленные частичным блокированием каждого из 5 ферментов, катализирующих в печени реакции синтеза мочевины, а также N-ацетилглутаматсинтазы. Эти генетические дефекты, очевидно, являются частичными. Полное блокирование какой-либо из стадий цикла мочевины в печени, по-видимому, несовместимо с жизнью, потому что другого эффективного пути удаления аммиака не существует.
Общим признаком всех нарушений синтеза мочевины является повышенное содержание NH 4 + в крови (гипераммониемия). Наиболее тяжёлые клинические проявления наблюдаются при дефекте фермента карбамоилфосфатсинтетазы. Клиническими симптомами, общими для всех нарушений цикла мочевины, являются рвота, нарушение координации движений, раздражительность, сонливость и умственная отсталость. Если заболевание не диагностируется, то быстро наступает гибель. У детей старшего возраста проявлениями заболевания служат повышенная возбудимость, увеличение размеров печени и отвращение к пище с высоким содержанием белка.
Лабораторная диагностика заболеваний включает определение содержания аммиака и метаболитов орнитинового цикла в крови, моче и спинномозговой жидкости; в сложных случаях прибегают к биопсии печени.
Значительное улучшение наблюдается при ограничении белка в диете, при этом могут быть предотвращены многие нарушения мозговой деятельности. Малобелковая диета приводит к снижению содержания аммиака в крови и к улучшению клинической картины при мягких формах этих наследственных нарушений. Пищу следует принимать часто, небольшими порциями, для того чтобы избежать резкого повышения уровня аммиака в крови.
24.4.5. Клинико-диагностическое значение определения мочевины в крови и моче. В крови здорового человека содержание мочевины составляет 3,33 – 8,32 ммоль/л. За сутки с мочой выводится 20 – 35 г мочевины.
Изменения содержания мочевины в крови при заболеваниях зависят от соотношения процессов её образования в печени и выведения почками. Повышение содержания мочевины в крови (гиперазотемия) отмечается при почечной недостаточности, снижение – при недостаточности печени, при диете с низким содержанием белков.
Повышение экскреции мочевины с мочой наблюдается при употреблении пищи с высоким содержанием белков, при заболеваниях, сопровождающихся усилением катаболизма белков в тканях, при приёме некоторых лекарств (например, салицилатов). Снижение экскреции мочевины с мочой характерно для заболеваний и токсических поражений печени, заболеваний почек, сопровождающихся нарушением их фильтрационной способности.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом