
Метаболизм триацилглицеридов (ТАГ). Синтез высших жирных кислот (ВЖК). Тканевой липолиз. Регуляция. Окисление глицерина. Бета-окисление ВЖК. Значение. Кетогенез. Локализация синтеза. Источники жирных кислот для синтеза. Образование фосфатидной кислоты.
| Рубрика | Химия |
| Предмет | Химия |
| Вид | презентация |
| Язык | русский |
| Прислал(а) | incognito |
| Дата добавления | 25.12.2016 |
| Размер файла | 83,2 K |
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Подобные документы
Химические, физические свойства жирных кислот. Способы производства жирных кислот: окисление парафинов кислородом воздуха; окисление альдегидов оксосинтеза кислородом. Гидрокарбоксилирование олефинов в присутствии кислот. Жидкофазное окисление олефинов.
контрольная работа [45,5 K], добавлен 15.03.2010
Высшие жирные кислоты. Биосинтез карбоновых кислот. Сложные эфиры высших одноатомных спиртов и высших жирных кислот. Простые липиды триацилглицерины. Реакции окисления липидов с участием двойных связей. Окисление с расщеплением углеводородного скелета.
реферат [1,0 M], добавлен 19.08.2013
Жиры как природные органические соединения, полные сложные эфиры глицерина и одноосновных жирных кислот. Лецитины как сложные эфиры глицерина, фосфорной и жирных кислот. Структурная формуладипальмитоилфосфатидихолина. Значение кардиолипина в медицине.
реферат [137,9 K], добавлен 10.06.2015
Аэробное окисление углеводов — основной путь образования энергии для организма. Клеточное дыхание — ферментативный процесс, результате которого, молекулы углеводов, жирных кислот и аминокислот расщепляются, освобождается биологически полезная энергия.
реферат [20,9 K], добавлен 17.01.2009
Обработка семян спиртовым раствором щелочи при повышенных температурах. Сложные эфиры трехатомного спирта глицерина и жирных кислот. Запасные и структурные липиды. Жирорастворимые витамины и защитные липиды. Продукты неполного синтеза и гидролиза липидов.
контрольная работа [49,9 K], добавлен 21.10.2013
7.11. Обмен триацилглицеринов
7.11.1. Распад триацилглицеринов в тканях (липолиз)
Триацилглицерины поэтапно расщепляется тканевыми липазами.

Ключевым ферментом липолиза является гормональнозависимая ТАГ-липаза. Образующиеся на этом этапе распада жиров глицерин и жирные кислоты окисляются в тканях с образованием энергии.
7.11.1.1. Окисление жирных кислот.
Различают несколько вариантов окисления жирных кислот: α — окисление, β — окисление, ω — окисление. Основным вариантом окисления жирных кислот является β — окисление. Оно наиболее активно протекает в жировой ткани, печени, почках и сердечной мышце.
β — окислениезаключается в постепенном отщеплении от жирной кислоты двух углеродных атомов в виде ацетил — КоА с освобождением энергии. Запас жирных кислот в клетках сосредоточен в цитозоле, где протекает активация жирных кислот с образованием ацил — КоА
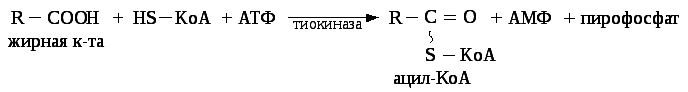
Сам процесс β-окисления ацил-КоА происходит в митохондриях. Митохондриальная мембрана непроницаема для длинноцепочечных ацил — КоА. В переносе их внутрь митохондрий участвует специальный переносчик карнитин(метил, гидропроизводное аминомасляной кислоты). Ацил — КоА образует с карнитином комплекс, который после переноса жирной кислоты внутрь митохондрий распадается.
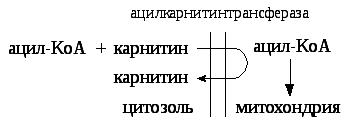
Энергетическая эффективность бета — окисления жирных кислот складывается из энергии окисления ацетил — КоА в цикле Кребса и энергии, освобождающейся в самом бета – цикле. Энергия окисления жирной кислоты тем выше, чем длиннее её углеродная цепь. Количество молекул ацетил — КоА из насыщенной жирной кислоты и количество образующихся из них молекул АТФ определяется по формулам:
n=N/2, гдеn-количество молекул ацетил — КоА,N- число атомов углерода в жирной кислоте.
Количество молекул АТФза счёт окисления молекул ацетил-КоА= (N/2)*12
Число β — циклов окисления на один меньше, чем количество образующихся молекул ацетил-КоА, поскольку в последнем цикле масляная кислота за один цикл переходит в две молекулы ацетил-КоА, и рассчитывается по формуле
Количество молекул АТФв β — цикле рассчитывается, исходя из последующего окисления образовавшихся в нём НАДН2(3 АТФ) и ФАДН2 (2 АТФ) по формуле
Количество молекул АТФ, образующихся в β — циклах= ((N/2)-1)*5
2 макроэргические связи АТФ расходуются на активацию жирной кислоты
Суммарная формуладля подсчёта выхода АТФ при окислении насыщенной жирной кислоты имеет вид:17(N/2)-7.
Окисление ненасыщенных жирных кислотна начальных стадиях представляет обычное β — окисление до места двойной связи. Если эта двойная связь находится в β — положении, то продолжается окисление жирной кислоты со второго этапа (минуя стадию восстановления ФАД→ ФАДН2). Если двойная связь находится не β — положении, то ферментами еноилтрансферазами связь перемещается в β – положение. Таким образом, при окислении ненасыщенных жирных кислот образуется меньше энергии (теряется образование ФАДН2 на каждую двойную связь). Она рассчитывается по формуле:
Химизм β — окисления насыщенных жирных кислот
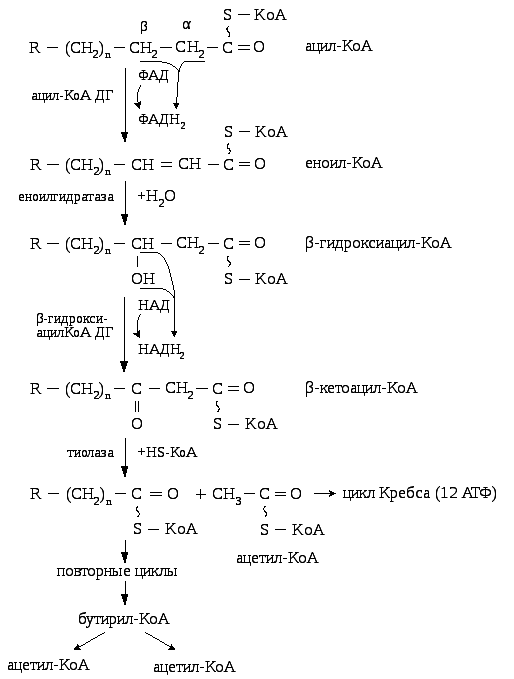
7.11.1.2. Окисление глицерина
На первом этапе глицерин активируется в глицерофосфат, затем окисляется до фосфодигидроксиацетона, который через стадию образования пирувата переходит в ацетил – КоА и окисляется в цикле Кребса.

7.11.2. Синтез триацилглицеринов (липогенез)
Для синтеза ТАГ необходимы глицерин и жирные кислоты.
Если глицерин поступает с пищей в недостаточном количестве, он может синтезироваться из углеводов через стадию образования общего метаболита фосфодигидроксиацетона.
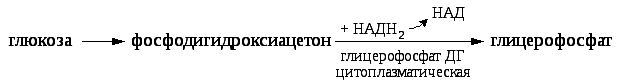
Активная форма глицерина – глицерофосфат используется на синтез ТАГ и глицерофосфолипидов.
7.11.2.2. Синтез жирных кислот
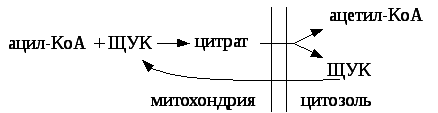
Синтез насыщенных жирных кислот происходит в цитозоле при участии сложного полиферментного комплекса (синтетаза жирных кислот или пальмататсинтетаза). Этот комплекс включает в себя особый ацилпереносящий белок и 6 ферментов. Для синтеза жирных кислот донором водорода является НАДФН2, образующийся в пентозофосфатном пути окисления глюкозы. Исходным веществом для синтеза жирных кислот является ацетил — КоА, который образуется в митохондриях в результате бета – окисления жирных кислот, и самостоятельно выйти в цитозоль не может. Транспорт ацетил — КоА из митохондрий в цитозоль осуществляется с помощью цитратного челночного механизма:
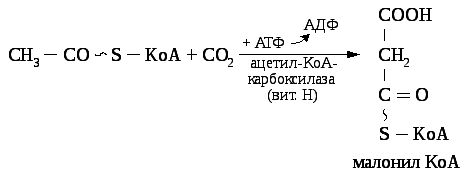
Ацетил — КоА на первом этапе карбоксилируется в малонил — КоА
Затем ацетил — КоА и малонил — КоА соединяются с полиферментным комплексом (синтетаза жирных кислот), в котором имеется две SН-группы, принадлежащие ацилпереносящему белку.
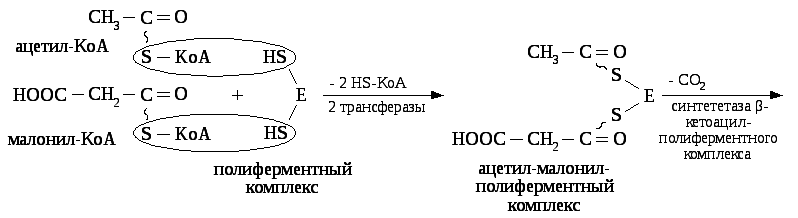
Затем остаток ацетила переносится на остаток малонила с образованием β – кетоацил — полиферментного комплекса и освобождением одной SH– группы полиферментного комплекса.
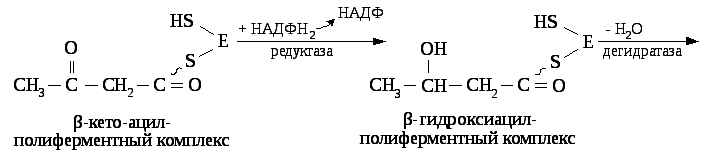
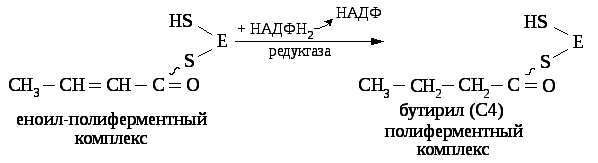
В последующем к этому комплексу с четырёх углеродной масляной кислотой присоединяется новая молекула малонил — КоА и в результате синтезируется капроновая кислота (С6) и далее до С16(пальмитиновая кислота). Более длинные жирные кислоты синтезируется с участием дополнительных ферментов элонгаз.
В тканях организма человека синтезируется только мононенасыщенные кислоты:
олеиновая и пальмитолеиновая. Они образуются из соответствующих насыщенных жирных кислот при участии ферментов десатураз (монооксигеназ) и цитохрома Р450.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
3.1. Обмен триглицеридов в тканях
Триглицериды синтезируются в клетках различных органов и
тканей в качестве резервных питательных веществ, однако их син
тез с наибольшей интенсивностью протекает в клетках печени и в
клетках жировой ткани. Для синтеза необходимы высшие жирные
кислоты и глицерол. Высшие жирные кислоты или поступают в клет
ки из плазмы крови, или же синтезируются в них из ацетилКоА.
Глицерол может поступать в клетки из плазмы крови, однако ос
новным источником глицерола для синтеза триглицеридов и фосфо
липидов в клетках служит фосфодигидроксиацетон промежуточный
продукт расщепления глюкозы.
Высшие жирные кислоты участвуют в биосинтезе триглицери
дов в виде своих активированных производных ацилКоА. Необ
ходимый для синтеза 3фосфоглицерол образуется или путем вос
становления фосфодигидроксиацетона ( реакция катализируется
глицеральдегид3фосфатдегидрогеназой за счет обратимости ее
действия ), или за счет фосфорилирования свободного глицерола
(реакция катализируется АТФзависимой глицеролкиназой).
Схема метаболического пути биосинтеза триглицеридов
После образования 3фосфоглицерола за счет двух последо
вательных реакций ацилирования образуется фосфатидная кислота.
От нее гидролитическим путем отщепляется остаток фосфорной
кислоты с образованием диглицерида, а затем с помощью еще од
ной реакции ацилирования завершается синтез триацилглицерина.
Синтез резервных триацилглицеринов идет в основном в пе
риод абсорбции продуктов пищеварения и поступления их во внут
реннюю среду организма. В постабсорбционном периоде идет моби
лизация резервных триглицеридов. Они расщепляются в клетках
под действием ферментов липаз.
При распаде триглицеридов в липоцитах жировой ткани по
последним данным работают три различных фермента по схеме:
Наименьшей активностью среди трех ферментов, участвующих
в расщеплении триацилглицерина обладает триацилглицеридлипаза,
поэтому активностью именно этого фермента определяется ско
рость гидролиза триглицеридов в целом. Триацилглицероллипаза
является регуляторным ферментом, активность которого изменяет
ся под влиянием ряда гормонов, таких как норадреналин, адрена
3.2. Обмен фосфолипидов в тканях
Все необходимые организму глицерофосфолипиды могут синте
зироваться в его клетках, причем в клетках могут функциониро
вать несколько альтернативных метаболических путей биосинтеза
При наличии в клетках свободных аминоспиртов может функ
ционировать тот же самый путь биосинтеза этаноламинфосфатидов
или холинфосфатидов, что и при их ресинтезе в стенке кишечни
ка. Поскольку мы его ранее уже рассматривали, представлена
лишь краткая схема этого метаболического пути:
Альтернативным вариантом синтеза может быть путь синтеза
с промежуточным образованием активированной формы фосфатидной
кислоты. Сама фосфатидная кислота образуется уже известным нам
путем, а далее она взаимодействует с цитидинтрифосфатом (ЦТФ)
с образованием активного фосфатидата. Этот вариант синтеза ра
ботает в клетках в тех случаях, когда в них нет свободных ами
носпиртов. Серин же является заменимой аминокислотой и может
синтезироваться из 3фосфоглицерата промежуточного продукта
расщепления глюкозы. Разумеется, синтез серина будет возмож
ным лишь при одновременном наличии в клетке источника аминного
Схема альтернативного пути синтеза
Синтезированный таким образом фосфатидилсерин может быть
преобразован в клетке в фосфатидилэтаноламин и, далее, в фосфа
Рассматриваемый вариант синтеза играет важную роль в ме
таболизме клеток еще и потому, что с его помощью в клетках
синтезируются инозитолфосфатиды, которые, как мы узнаем нес
колько позднее, играют существенную роль в механизме действия
Кроме ранее рассмотренных путей фосфатидилсерин может
быть синтезирован в клетке в ходе обменной реакции:
Расщепление глицерофосфолипидов в клетках идет при учас
тии ферментов фосфолипаз. Фосфолипаза А2 катализирует гидролиз
сложноэфирной связи между ацильным остатком и вторым атомом
углерода глицерола. Образовавшийся лизофосфолипид может либо
подвергаться реацилированию с образованием новой молекулы гли
церофосфолипида, либо при участии фермента лизофосфолипазы те
ряет второй ацильный остаток, превращаясь таким образом в гли
церолфосфорильное производное. Последнее в свою очередь может
расщепляться гидролазой до глицерол3фосфата и аминоспирта.
Возможен альтернативный путь расщепления глицерофосфоли
пидов, в ходе которого вначале под действием фосфолипазы А1 от
фосфолипида отщепляется ацильный остаток от «C1» глицерола,
затем под действием фосфолипазы А2 отщепляется второй ацильный
остаток и на заключительном этапе идет отщепление аминоспирта
с образованием свободного глицерол3фосфата.
3.3. Представление о путях синтеза и распада сфинголипидов
Сфинголипиды, подобно глицерофосфолипидам, не являются незаменимыми компонентами пищи и могут синтезироваться из других соединений. Для их синтеза нужен в первую очередь сфингозин, который образуется в ходе нескольких последовательных реакций из пальмитоилКоА и серина; необходимы активированные жирные кислоты в виде ацилКоАпроизводных; необходимы также или активированный холин в виде ЦДФхолина для синтеза сфинго
Читайте также: Ткань дерева 6 букв четвертая э
миелинов, или активированные мономеры углеводной природы в ви
де их УДФпроизводных для синтеза цереброзидов или ганглиози
Принципиальная схема синтеза сфинголипидов:
При синтезе ганглиозидов активированной формой сиаловой кис
лоты является ее ЦДФпроизводное.
Расщепление сфинголипидов в клетках происходит в лизосо
мах при участии имеющихся в этих органеллах различных кислых
лизосомальных гидролаз. Углеводные компоненты гликосфинголипи
дов расщепляются при участии различных лизосомальных гликози
даз. Сфингомиелин расщепляется на церамид и фосфорилхолин при
участии сфингомиелиназы. Образовавшийся же при деградации раз
личных сфинголипидов церамид гидролизуется при участии церами
дазы на сфингозин и высшую жирную кислоту. Продукты деградации
поступают из лизосом в цитозоль, где они могут использоваться
в биосинтезах или расщепляться до конечных продуктов.
4.1. Интеграция метаболических путей обмена липидов
Ранее нами были рассмотрены отдельные метаболические пути,обеспечивающие синтез и расщепление молекул липидов различных классов. В клетке эти метаболические пути взаимодействуют друг с
другом, обеспечивая переключение потока вещества с одного метабо
лического пути на другой в соответствии с потребностями клеток.
Кроме того, в любой живой системе обмен липидов связан с обменом
соединений других классов,например, обменом углеводов или амино
кислот. Эти взаимосвязи можно проследить, воспользовавшись общей
Из схемы следует, что пищевые липиды являются источниками
высших жирных кислот, глицерола, аминоспиртов и некоторых других
соединений, используемых организмом для синтеза свойственных для
него структурных или резервных липидов. Свободные высшие жирные
кислоты, наряду с глицеролом и аминоспиртами образуются в орга
низме также при расщеплении резервных или структурных липидов. Еще одним источником высших жирных кислот может служить их синтез
из ацетилаКоА, который в свою очередь может быть промежуточным
продуктом обмена углеводов или аминокислот. Заметим, что жирные
кислоты в клетках используются в различных метаболических путях
клетки только в их активированной форме в виде ацилКоА.
Одним из ключевых метаболитов липидного обмена является аце
тилКоА, поскольку, вопервых, именно через это соединение осу
ществляется окислительное расщепление высших жирных кислот;
вовторых, через ацетилКоА атомы углерода жирных кислот могут
быть использованы для пластических целей для синтеза холестеро
ла или полипреноидов; в третьих, через ацетилКоА в гепатоцитах
углеродные цепи жирных кислот преобразуются в ацетоновые тела
гидрофильные «топливные» молекулы, легко транспортируемые в клет
ки различных органов и тканей; в четвертых, через ацетилКоА осу
ществляются метаболические превращения углеродных скелетов амино
кислот и моносахаридов в жирные кислоты, используемые в дальней
шем для синтеза более сложных липидных молекул.
Соединения других классов аминокислоты и моносахариды в
ходе своего метаболизма образуют промежуточные продукты, которые
могут в дальнейшем использоваться в клетке как для синтеза высших
жирных кислот, так и для образования других мономерных единиц,
необходимых для синтеза сложных липидов: глицерола, этаноламина,
холина, сфингозина и пр. Таким образом, обмен липидов оказывается
тесно связанным с обменом соединений других классов, а метаболи
ческие пути обмена липидов различных классов являются частью об
щей метаболической сети, функционирующей в организме.
4.2. Регуляция обмена липидов на уровне организма
Липиды выполняют множество функций в организме, одной из важнейших среди них является обеспечение клеток различных органов и тканей энергией, т.к. от 30% до 40% всей необходимой ему энергии человек получает за счет окислительного расщепления соединений липидной природы. Интенсивность и направленность различных
превращений липидов должны соответствовать потребности организма в энергетическом и пластическом материале. Поэтому крайне важными становятся как вопросы регуляции обмена липидов на уровне организма, так и проблема координации функционирования метаболических путей обмена липидов и метаболических путей обмена соединений
других классов, обеспечивающих снабжение клеток необходимой им
энергией. В конечном итоге, эффективная работа регуляторных и ко
ординирующих механизмов обеспечивает адаптацию организма к изме
няющимся условиям его существования.
Так, в постабсорбционном периоде, когда поступление глюкозы
и экзогенных липидов из кишечника во внутреннюю среду организма
прекращается, потребность организма в энергии покрывается за счет
расщепления резервных триглицеридов, основная масса которых сос
редоточена в жировой ткани. В ходе мобилизация резервных триглице
ридов образуются высшие жирные кислоты и глицерол, которые посту
пают из липоцитов вначале в кровь, а затем в клетки различных ор
ганов и тканей, где и окисляются с выделением необходимой клеткам
Этот процесс мобилизация резервных триглицеридов или ли
полиз стимулируется рядом гормонов, к числу которых относятся ад
реналин, норадреналин, глюкагон, bлипотропный гормон гипофиза,
соматотропин, АКТГ, МСГ, кортизол, тироксин, тестостерон. Многие
из этих гормонов являются активаторами гормончувствительной ли
пазы липоцитов (триацилглицероллипазы). Для оптимального протека
ния большинства липолитических процессов необходимо присутствие
кортизола, соматотропина и гормонов щитовидной железы. Сами по
себе эти гормоны не оказывают прямого влияния на липолиз, а дейс
твуют как факторы, стимулирующие действие других гормонов.
Важнейшая роль в мобилизации резервных липидов в организме
человека принадлежит адреналину ( вместе с норадреналином ), ко
торый выделяется в жировой ткани нервными окончаниями симпатичес
кой нервной системы. Вторым источником адреналина является мозго
вое вещество надпочечников, откуда адреналин доставляется в жиро
вую ткань с током крови. Вероятно, адреналин из мозгового вещест
ва надпочечников играет важную роль в мобилизации триглицеридов
жировой ткани в условиях острого эмоционального стресса. Механизм
активации липолиза при воздействии на липоциты адреналина предс
РУСЛО КРОВИ ЦИТОЗОЛЬ ЛИПОЦИТА
Адреналин взаимодействует со своим рецептором на наружной по
верхности мембраны липоцита с образованием гормонрецепторного
комплекса. В ответ на образование гормонрецепторного комплекса с
помощью специального механизма происходит активация расположенной
на внутренней поверхности наружной клеточной мембраны липоцита
аденилатциклазы фермента, синтезирующего из АТФ циклическую АМФ
(цАМФ). Увеличение внутриклеточной концентрации цАМФ активирует
фермент протеинкиназу, которая осуществляет активацию триацилгли
цероллипазы путем ее фосфорилирования, т.е. путем ковалентной мо
дификации фермента. Поскольку скорость липолиза лимитируется
активностью триацилглицероллипазы, активация фермента приводит к
ускорению гидролиза резервных триглицеридов и увеличению выхода
высших жирных кислот и глицерола из липоцита в русло крови.
Гормоны глюкагон, bлипотропин, меланоцитстимулирующий гормон, кортикотропин активируют липолиз в жировой ткани, увеличивая концентрацию цАМФ в липоцитах с помощью механизма, сходного с механизмом активации липолиза под действием адреналина. Интересно,
что существуют видовые различия в эффективности функционирования
этих регуляторных механизмов: так, у птиц глюкагон является мощным
стимулятором липолиза, тогда как липолитический эффект глюкагона
у человека крайне незначителен.
Соматотропный гормон не оказывает прямого влияния на ско
рость расщепления триглицеридов в липоцитах, однако соматотропин
увеличивает скорость синтеза аденилатциклазы за счет ускорения
процесса транскрипции соответствующего гена. Увеличение содержа
ния аденилатциклазы в липоцитах увеличивает эффект воздействия на
жировую ткань таких гормонов как адреналин, bлипотропин и др.
Сходным образом оказывает стимулирующее влияние на липолиз и
кортизол, поскольку этот гормон увеличивает содержание в липоци
тах другого фермента гормончувствительной липазы. Кортизол
выступает в качестве стимулятора транскрипции гена, ответственно
го за синтез этого фермента. Повышение же содержания триацилгли
цероллипазы в липоцитах способствует более быстрому и более выра
женному ответу клеток на воздействие на них гормонов типа адрена
Механизм действия тироксина на жировую ткань не совсем ясен.
Известно, что этот гормон способствует более эффективной передаче
стимулирующего сигнала с гормонрецепторного комплекса на адени
латциклазу, в результате чего при воздействии на липоциты гормо
нов типа адреналина происходит более быстрая активация липолиза в
Основным гормоном, тормозящим липолиз в жировой ткани, явля
ется инсулин. Инсулин снижает содержание цАМФ в липоцитах, пови
димому, за счет активации фосфодиэстеразы, переводящей цАМФ в
обычную АМФ. Снижение концентрации цАМФ в клетках приводит как к
инактивации протеинкиназы, так и к активации фосфопротеинфосфата
зы, в результате чего происходит дефосфорилирование гормончувс
твительной липазы с ее инактивацией и последующим торможением ли
полиза. Простагландины также снижают содержание цАМФ в липоцитах
с последующим торможением липолиза в клетках.
В период абсорбции в клетках различных органов и тканей активно идет липогенез. Во внутреннюю среду организма из кишечника поступают глюкоза и другие моносахариды, а также триацилглицерины в составе ХМ или ЛПОНП. Моносахариды, поступающие в липоциты или в гепатоциты, используются в ходе липогенеза, являясь как источ
никами ацетилКоА для синтеза высших жирных кислот, так и источ
никами фосфотриоз, необходимых для образования 3фосфоглицерола.
Триглицериды ХМ или ЛПОНП после их гидролиза липопротеидлипазой
также являются источниками высших жирных кислот и глицерола, пос
тупающих в клетки и в дальнейшем используемыми в качестве субс
Гормоном, стимулирующим липогенез, является инсулин. Инсулин
ускоряет поступление глюкозы в клетки и стимулирует ее фосфорили
рование, запуская тем самым процесс утилизации глюкозы в клетках.
Причем стимулируется как процесс аэробного окисления глюкозы до
СО2 и Н2О, так и работа пентозного цикла окисления глюкозы, обес
печивающего клетки восстановительными эквивалентами в виде
Инсулин активирует работу пируватдегидрогеназного комплекса, что приводит к увеличению образования ацетилКоА исходного субстрата для синтеза высших жирных кислот. Инсулин повышает активность фермента ацетилКоАкарбоксилазы, катализирующего превращение ацетилКоА в малонилКоА, также необходимого для синтеза высших жирных кислот. Ускорение окислительного распада глюкозы в клетке приводит также к увеличению в ней концентрации фосфотриоз 3фосфоглицеринового альдегида и фосфодигидроксиацетона, ис
пользуемых для образования 3фосфоглицерола. Таким образом, воз
действие инсулина на клетки приводит к наработке в них исходных
соединений для синтеза триглицеридов. Кроме того, инсулин активи
рует в клетках глицеролфосфатацилтрансферазу фермент, катали
зирующий перенос ацильного остатка с КоА на 3фосфоглицерол
первую реакцию метаболического пути синтеза триацилглицеринов.
Регуляция активности пируватдегидрогеназного комплекса, аце
тилКоАкарбоксилазы и глицеролфосфатацилтрансферазы осуществля
ется путем координированного процесса ковалентной модификации
этих ферментов ( фосфорилирование дефосфорилирование ).
В целом, воздействие инсулина на липоциты приводит, вопер
вых, к торможению липолиза в клетках, а, вовторых, к активации в
них процесса липогенеза, способствуя тем самым накоплению энерге
тических резервов в организме в виде триацилглицеринов.
4.3.Интеграция и регуляция обмена глюкозы и высших жирных кислот на клеточном уровне
Известно, что в постабсорбционном состоянии основным энерге
тическим «топливом» для клеток являются высшие жирные кислоты,
тогда как в период пищеварения, когда во внутреннюю среду орга
низма поступают моносахариды и ресинтезированные в стенке кишеч
ника триглицериды, основным энергетическим топливом становится
глюкоза; более того, поступающая в клетки глюкоза превращается в
жирные кислоты. Последний процесс особенно характерен для гепато
При поступлении глюкозы в клетки она в цитозоле окисляется
до пирувата (см. следующую далее схему), последний проходит через
внутреннюю мембрану митохондрий и окисляется в матриксе до аце
тилКоА. Образовавшийся ацетилКоА конденсируется с оксалоацета
том (ЩУК) с образованием цитрата, а цитрат выходит из митохондрии
Поступивший в цитозоль цитрат, вопервых, служит источником
ацетилКоА и восстановительных эквивалентов для синтеза высших
жирных кислот, а, вовторых, активирует фермент ацетилКоАкар
боксилазу, стимулируя тем самым образование малонилКоА, также не
обходимого для синтеза высших жирных кислот. В результате при из
Читайте также: Хохлома роспись по ткани
бытке глюкозы в клетке запускается синтез жирных кислот.
МалонилКоА в свою очередь угнетает перенос высших жирных кислот из цитозоля в матрикс митохондрий, ингибируя активность внешней ацетилКоА:карнитинацилтрансферазы, выключая таким образом окисление высших жирных кислот
В итоге при поступлении глюкозы в клетку угнетается окисле
ние высших жирных кислот, стимулируется их синтез, а потребность
клетки в энергии покрывается за счет аэробного окисления глюкозы,
чему способствует повышение концентрации ацетилКоА и цитрата в
матриксе митохондрий. Увеличение концентрации жирных кислот в
клетке наряду с нарастанием концентрации в них триозофосфатав
создает условия для синтеза резервных триглециридов. В этот про
цесс включаются также высшие жирные кислоты и глицерол, поступаю
щие в клетку в результате гидролиза триглицеридов ХМ и ЛПОНП.
В постабсорбционном периоде, когда концентрация глюкозы в клетках снижается, поток цитрата из митохондрий в цитозоль уменьшается, в результате в цитозоле уменьшается концентрация ацетилКоА и инактивируется ацетилКоАкарбоксилаза. Снижается содержание малонилКоА, что приводит как к прекращению синтеза высших жирных кислот, так и к снятию ингибирования ацилКоА:карнитинацилтрансферазы и восстановления транспорта жирных кислот в
матрикс митохондрий, где они начинают окисляться. Таким образом,
в условиях недостатка глюкозы в клетках выключается синтез высших
жирных кислот и включается их bокисление, которое и становится
основным источником свободной энергии в клетках.
4.4.Патология липидного обмена
Нарушения липидного обмена выявляются у людей с самыми раз
личными заболеваниями. Эти нарушения можно разделить на первичные
и вторичные. При первичных или наследственных нарушениях липид
ного обмена патологические состояния возникают как следствие гене
тического дефекта, сопровождающегося нарушением синтеза белковых
молекул, имеющих то или иное отношение к обмену липидов. Это мо
жет быть нарушение синтеза белковрецепторов для ЛПНП, или нару
шение синтеза апопротеинов, или, наконец, нарушение синтеза фер
ментов, катализирующих отдельные реакции липидного обмена.
Вторичные нарушения липидного обмена развиваются или как
следствие имеющегося заболевания, например, сахарный диабет, или
как следствие воздействия факторов внешней среды, включая сюда и
нарушение поведенческих реакций. Примерами могут служить наруше
ния обмена липидов при отравлении четыреххлористым углеродом или
ожирение при систематическом переедании.
4.1. Первичные нарушения обмена липидов
К наследственным заболеваниям, сопровождающимся нарушениями
обмена липидов относятся, например, гиперхиломикронемия, семейная
гиперхолестеринемия, болезнь НиманаПика, болезнь ТеяСакса и ряд
других патологических состояний.
4.1.1. Наследственная гиперхиломикронемия
При наследственной гиперхиломикронемии у больных нарушена функция фермента липопротеидлипазы в результате или нарушения образования самого фермента, или в результате нарушения синтеза апопротеина СII, являющегося активатором липопротеидлипазы. В крови вследствие ингибирования расщепления триглицеридов накапливаются хиломикроны и липопротеиды очень низкой плотности. В крови
даже натощак повышено содержание триглицеридов. У таких больных
развивается гепатоспленомегалия, часты боли в животе, часты панк
реатиты. Для этих больных характерны ксантомы доброкачественные
опухоли из подкожной жировой ткани.
При этом заболевании в организме нарушен синтез рецепторов
для ЛПНП, в результате чего нарушена утилизация этих липопротеи
дов. В крови таких больных повышено содержание ЛПНП и холестеро
ла. Содержание холестерола в крови может в несколько раз превы
шать норму. Это накопление в крови ЛПНП и холестерола быстро, уже
в юношеском возрасте, приводит к развитию атеросклероза. Тяжесть
заболевания в значительной мере зависит от того, один или оба ге
на белковрецепторов ЛПНП дефектны. При дефекте одного из генов в
клетках имеется половинное количество рецепторов для ЛПНП, если
дефектны оба гена рецепторов для ЛПНП вообще нет. Без соответс
твующего лечения больные редко достигают 30летнего возраста, они
обычно погибают от инфаркта миокарда.
При болезни НиманаПика в клетках больного отсутствует фер
мент лизосом сфингомиелиназа или же его активность значительно
снижена. В лизосомах накапливается сфингомиелин, т.е. речь идет о
типичном варианте лизосомных болезней накопления. Поражаются се
лезенка, печень, мозг, почки и др. органы. Для больных характерна
задержка умственно и физического развития, нарушения функций раз
личных органов. Последствия ранняя смерть.
Болезнь ТеяСакса является еще одним примером наследственно
го нарушения обмена сфинголипидов. У больных, страдающих данным
заболеванием, в лизосомах отсутствует фермент Nацетилгексозамини
даза, в результате чего нарушается расщеплением ганглиозидов.
Особенно много ганглиозидов накапливается в лизосомах клеток моз
га. Для таких больных также характерна задержка умственного и фи
зического развития и смерть обычно в возрасте до 5 лет. Специфи
ческим признаком этого заболевания является ранняя слепота.
Частота врожденных нарушений обмена липидов широко варьиру
ет. Так, семейная гиперхолестеринемия встречается с средней час
тотой 1:200, тогда как болезнь ТеяСакса 1:300 000.
4.2.Вторичные нарушения обмена липидов
Из вторичных нарушений обмена липидов мы остановимся на жи
ровой дистрофии печени, ожирении, желчнокаменной болезни и ате
4.2.1. Жировая дистрофия печени
Сущность этого патологического процесса состоит в том, что в
гепатоцитах накапливаются липиды, причем преимущественно тригли
цериды. Масса триглицеридов в тяжелых случаях может составлять до
50% от массы печени. Естественно, гепатоциты, переполненные три
глицеридами, в конце концов погибают и замещаются фиброзной соеди
нительной тканью; развивается цирроз печени с нарушениями функций
органа. Ситуация может быть настолько тяжелой, что больные поги
бают в результате печеночной недостаточности в течение нескольких
суток это так называя острая желтая дистрофия печени.
Жировая дистрофия печени не является какимлибо специфичес
ким процессом. Она развивается в ответ на острую или хроническую
интоксикацию экзогенного или эндогенного происхождения. Так. жи
ровая дистрофия печени наблюдается при отравлениях некоторыми хи
мическими соединениями ( например, четыреххлористым углеродом ),
при отравлении некоторыми видами грибов, при алкоголизме, при тя
желом сахарном диабете, при туберкулезе и др.
Повидимому, в развитии жировой инфильтрации печени может быть задействовано несколько факторов. Вопервых, она может быть результатом увеличения содержания свободных высших жирных кислот в плазме крови, обусловленного или чрезмерным уровнем мобилиза ции жиров из жировых депо, или усиленным гидролизом триглицеридов
ХМ и ЛПОНП внепеченочной лиопротеидлипазой. В результате возрастает поглощение и эстерификация высших жирных кислот клетками пе
чени. Образующихся в печени ЛПОНП становится недостаточно для
эвакуации синтезированных в гепатоцитах триглицеридов и они на
капливаются в печени, вызывая ее жировое перерождение. Такова
причина развития жировой дистрофии печени, например, при тяжелом
сахарном диабете или при длительном потреблении пищи, содержащей
избыточное количество жира.
Вовторых, жировая дистрофия печени может быть обусловлена
нарушением образования в гепатоцитах липопротеидов, обеспечиваю
щих в норме эвакуацию триглицеридов из печени. В свою очередь,
нарушение образования липопротеидов в гепатоцитах может быть выз
вано: а) снижением синтеза апобелков, необходимых для формирова
ния липопротеидов; б) недостаточным поступлением или снижением
синтеза фосфолипидов, необходимых для формирования липопротеидных
частиц, в) нарушением формирования липопротеидов из апобелков и
фосфолипидов или нарушением работы механизма их экскреции.
Жировая дистрофия печени, наблюдающаяся при голодании, при
недостатке в пище незаменимых аминокислот, наконец, при алкого
лизме, обусловлена нарушением синтеза апобелков, необходимых для
формирования ЛПОНП и эвакуации триглицеридов.
Механизмы синтеза апобелков и фосфолипидов более чувствитель
ны к воздействию токсических соединений, нежели синтез высших
жирных кислот и триглицеридов, поэтому при воздействии ряда ток
сических агентов ( четыреххлористый углерод, хлороформ, свинец,
мышьяк) и наблюдается избыточное накопление триглицеридов в гепа
тоцитах. Оротовая кислота также вызывает жировое перерождение пе
чени; считают, что под действием оротовой кислоты нарушается про
цесс гликозилирования липопротеидов в аппарате Гольджи и ингиби
рует их дальнейший переход из гепатоцитов в плазму крови.
Жировое перерождение печени может стимулироваться при акти
вации перекисного окисления в мембранах гепатоцитов, при недос
татке некоторых витаминов (пиридоксин или пантотеновая кислота),
а также при недостатке в пище холина или метионина.
Нарушение синтеза апобелков может быть по крайней мере облегчено дачей больному полноценного белкового питания, обеспечивающего его организм всеми необходимыми для синтеза апобелков аминокислотами. Учитывая, что до 60% фосфолипидов ЛПОНП составляет фосфатидилхолин, дача пострадавшему холина будет способствовать нормализации синтеза фосфатидилхолина в гепатоцитах. Того же
эффекта можно добиться путем дополнительного поступления в орга
низм больного аминокислоты метионина, служащего источником ме
тильных группировок при эндогенном синтезе холина. В то же время
дача больному лекарственных препаратов, являющихся акцепторами
метильных групп, таких как витамин В5 или препаратов гуанидинового
ряда, нежелательно, так как они будут тормозить эндогенный синтез
фосфатидилхолина. Соединения типа холина или метионина получили
название липотропных веществ, а соединения типа никотиновой кис
лоты или гуанидинов носят название антилипотропных веществ.
Определенный вклад в жировую инфильтрацию печени может вно
сить и снижение скорости окисления высших жирных кислот в гепато
цитах вследствие недостатка карнитина переносчика жирных кислот
через мембрану митохондрий. Недостаток карнитина может наблюдать
ся при дефиците источника метильных групп для его синтеза, а им,
как известно, является Sаденозилметионин. Соответственно, дача
метионина будет способствовать увеличению содержания карнитина в
клетках и ускорять окисление высших жирных кислот в клетках.
4.2.2.Нарушение обмена липидов при ожирении
Избыточное накопление липидов в организме получило название
ожирение. Диагноз ожирение ставят в том случае, когда масса тела
обследуемого превышает оптимальную на 20%. Расчет оптимальной
массы тела можно произвести по простейшей формуле:
Многочисленные более сложные формулы для расчета не вносят су
щественных корректив в величину оптимальной массы отклонения не
превышают 35%. По данным американских страховых компаний опти
мальная масса для человека , рост которого 170 см, составляет при
худощавом телосложении 68 кг, а при крепком 73 кг. Подсчитано,
что каждый кг излишней массы сокращает продолжительность жизни на
резервных триглицеридов в жировых депо. Ожирение может быть первичным, обусловленным алиментарноконституциональными факторами, или же вторичным, в последнем случае оно является следствием либо имеющейся патологии, например, следствием эндок
ринных расстройств, либо следствием поведенческой реакции ( при
Различают два типа ожирения, гиперцеллюлярный и гипертрофи
ческий. При гиперцеллюлярном ожирении в организме увеличивается
количество адипоцитов: если в норме их число составляет величину
порядка 26х109 клеток, то у людей с гиперцеллюлярным типом ожире
ния их число может быть больше в 23 раза. В таком случае даже
при нормальном содержании жира в каждом отдельном адипоците общая
масса резервного жира может значительно превышать норму. Этот тип
ожирения явно носит наследственный характер. Известно: если у ре
бенка один из родителей страдает ожирением, то вероятность разви
тия этой патологии у ребенка составляет около 40%; если же ожире
ние есть у обоих родителей, то вероятность развития ожирения у ре
бенка возрастает до 80%. Правда, следует учитывать и обычаи, су
ществующие в данной конкретной семье склонность к избыточному
употреблению пищу (ребенок берет пример с папы и мамы).
При гипертрофическом ожирении количество адипоцитов в орга
низме остается нормальным, но увеличивается содержание триглице
ридов в каждом отдельном адипоците. В норме в адипоците содержит
ся до 0,6 мкг на клетку, тогда как при ожирении оно может возрас
Читайте также: Страны лидеры по производству синтетических тканей
Как при гипертрофическом, так и при гиперцеллюлярном ожире
нии увеличение массы тела связано с накоплением избытка триглице
ридов в результате превышения калорийности пищи над энергозатра
тами; без этого превышения не реализуется никакая наследственная
предрасположенность. Однако следует заметить, что при увеличенном
количестве липоцитов в организме потенциальная возможность для
развития ожирения значительно выше, так же как выше и общая ре
зервная емкость жировых депо. При лечении больных с гиперцеллю
лярным ожирением возникает больше сложностей, потому что снижение
массы тела не сопровождается уменьшением числа липоцитов и сохра
няется высокая степень предрасположенности к повторному нараста
нию массы резервного жира.
В метаболизме адипоцитов больных ожирением возникают опре
деленные изменения; в частности установлено, что:
а) повышается способность адипоцитов утилизировать внутрикле
б) в адипоцитах ускоряются процессы синтеза высших жирных
кислот и триглицеридов стимуляция липогенеза;
в) в адипоцитах увеличивается активность липолитических фер
ментов, в связи с чем в адипоцитах ускоряется процесс обмена ре
г) понижается чувствительность адипоцитов к инсулину, что яв
ляется следствием снижения числа рецепторов для инсулина на на
ружной клеточной мембране переполненных триглицеридами адипоцитов;
д) сохраняется чувствительность адипоцитов к жиромобилизующе
му действию катехоламинов.
Для больных ожирением характерна гиперлипидемия, особенно вы
раженная при II III степени ожирения. В крови повышено содержа
ние ЛПОНП и ЛПНП, а, следовательно, повышено содержание и тригли
церидов и холестерола, что способствует раннему развитию атероск
Для таких больных характерна гиперинсулинемия, что связано с
снижением чувствительности адипоцитов к инсулину изза уменьшения
числа инсулиновых рецепторов на поверхности этих клеток. После
приема пищи поступающая в кровь глюкоза медленно проникает в ади
поциты, в результате чего ее концентрация в крови повышена дли
тельное время после приема пищи. В ответ на повышение концентра
ции глюкозы островковый аппарат поджелудочной железы выбрасывает
инсулин, но повышение его концентрации в крови почти не дает эф
фекта. В результате в крови одновременно повышена концентрация и
глюкозы, и инсулина, что создает «благоприятные» условия для раз
вития сахарного диабета. Практически у всех больных с II и в осо
бенности с III степенью ожирения регистрируется нарушение толе
У больных ожирением регистрируются и другие нарушения функ
ций. Так, у них обычно снижена секреция катехоламинов, что тормо
зит липолиз в липоцитах и способствует дальнейшему накоплению жи
ра в жировых депо; у них наблюдаются также расстройства водносо
левого обмена с нарушением функций почек и др.
При проведении профилактической работы среди населения мало
рекомендовать людям увеличение физической нагрузки типа «нужно
больше ходить или бегать», поскольку физическая нагрузка приводит
к увеличению аппетита и потреблению избыточного количества пищи.
Акцент в этой работе должен быть смещен на достижение сбалансиро
ванности калорийности пищевого рациона и энергозатрат, поэтому на
селение нужно научить хотя бы ориентировочно рассчитывать кало
рийность рациона и величину энергозатрат. Без этого все разговоры
о профилактике распространения ожирения на популяционном уровне
останутся лишь благими пожеланиями.
Желчнокаменная болезнь это довольно широко распространенное заболевание, особенно среди людей пожилого возраста. Оно связано с появлением в желчевыводящих путей твердых конкрементов или желчных камней, которые становятся причиной или нарушения оттока желчи из желчевыводящих путей, или причиной воспалительного процесса в желчевыводящих путях. Обычно в желчных камнях основная их масса приходится на холестерол и билирубин, хотя при химическом анализе в них может быть обнаружено множество различных соединений. Если в составе камня более 70% его массы приходится на холестерол, то они относятся к холестериновым камням. Холестериновые камни встречаются в 2/3 случаев этого заболевания.
Избыток холестерола выделяется из организма в основном с желчью. Холестерол плохо растворим в воде, в связи с чем он в норме содержится в желчи в составе мицелл, обеспечивающих его растворение. В состав мицелл желчи входят также желчные кислоты и фосфолипиды ( в основном это фосфатидилхолин ), именно они обеспечивают растворимость холестерола в водной фазе желчи. Холестерол, повидимому, секретируется гепатоцитами уже в мицеллярной форме, хотя, возможно, также формирование мицелл и в первичной желчи.
Желчь из печени поступает в желчный пузырь, где происходит ее концентрирование за счет всасывания в стенку пузыря части воды. Одновременно происходит и всасывание части желчных кислот, поэтому в пузырной желчи происходит увеличение относительной концентрации холестерола по сравнению с концентрацией желчных кислот. Если указанный процесс приводит к нарушению структуры мицелл, то создаются условия для перехода холестерола из мицеллярной,устойчивой в растворе формы,в жидкокристаллическую форму, которая в воде неустойчива. При прогрессировании этого процесса в дальнейшим происходит переход холестерола в твердокристаллическую форму, что и приводит к образованию холестериновых камней.
В ряде случаев желчь может генерировать кристаллы холестерола еще до ее поступления в желчный пузырь, что наблюдается при нарушении желчеобразования непосредственно в печени. Повидимому, это связано или с большим избытком холестерола, поступающего в желчь, или же с снижением объема синтеза желчных кислот. Способность желчи генерировать конкременты, в том числе и преимущественно холестериновой природы, получила название литогенности желчи ( от слова litos камень ).
Литогенность желчи может быть оценена с помощью различных методов исследования. При использовании биохимических методов исследования в желчи определяют содержание холестерола, желчных кислот ( холатов), иногда также определяют содержание фосфатидилхолина ( лецитина ). Далее рассчитывают холатно/холестериновый коэффициент, т.е. отношение концентраций желчных кислот и холестерола. У здорового человека значение холатнохолестеринового коэффициента больше 10. Если полученное значение коэффициента менее 10, желчь считается литогенной.
Более точно литогенность желчи можно определить, учитывая содержание в ней не только холатов и холестерола, но и лецитина. Одним из методов такой оценки является графический способ анализа результатов исследования с использованием треугольной системы координат ( так называемый «треугольник Myant»).
Химические методы исследования занимают сравнительно много времени. Если вопрос о литогенности желчи нужно решить срочно , например, во время операции, то можно воспользоваться методом поляризационной микроскопии. С помощью поляризационной микроскопии можно решить, находится ли холестерол в данной желчи только в составе мицелл, и тогда желчь нелитогенна. Или же наряду с мицеллярной формой в желчи холестерол присутствует также в жидкокристаллической ( неустойчивой ) форме, или в твердокристалличесской форме. В двух последних случаях желчь будет литогенной.
До настоящего времени основным методом лечения желчнокаменной болезни является хирургический. Это или тяжелая операция по удалению желчного пузыря, или же ультразвуковое дробление желчных камней в желчевыводящих путях. Однако начинает применяться и другой метод постепенное растворение камней с помощью длительного
приема хенодезоксихолевой кислоты, от содержания которой в желчи в значительной мере зависит растворимость в ней холестерола. Установлено, что ежедневный прием 1 г хенодезоксихолевой кислоты в течении года может привести к растворению холестеринового камня размером с горошину. Использование хенодезоксихолевой кислоты целесообразно еще и потому, что она оказывает ингибирующее действие на ГМГредуктазу в гепатоцитах, снижая тем самым уровень эндогенного синтеза холестерола в организме. Снижение эндогенного синтеза холестерола приводит к уменьшению его концентрации в желчи, что ведет к уменьшению ее литогенности.
Наиболее распространенным нарушением липидного обмена является атеросклероз. Это патологическое состояние связано с нарушениями в стенках крупных сосудов аорты или крупных артерий, вызываемыми избыточным накоплением в них холестерола. Проявлениями атеросклероза могут быть различные заболеваниями: ишемическая болезнь сердца ( стенокардия или инфаркт миокарда ), инсульт, гангрена конечности и др. Значимость проблемы атеросклероза можно проиллюстрировать следующим примером: в средине 80х годов в США на 220 млн населения регистрировалось 1,5 млн инфарктов и 550 тысяч смертных случаев от этого заболевания в год и в большинстве случаев причиной инфаркта было атеросклеротическое поражение сосудов. Механизм развития атеросклеротических процессов в сосудах еще полностью не выяснен. Не исключено, что атеросклероз может быть финалом развития достаточно разнородных процессов, однако огромное большинство ученых считает, что нарушения липидного обмена вносят существенный вклад в развитие этой патологии.
Атеросклеротические изменения в стенке сосудов начинаются с образования липидных пятен или полосок на внутренней поверхности аорты или крупных артерий. Они имеют желтоватую окраску и могут быть обнаружены даже у детей. Но эти изменения могут регрессировать, они не создают какихлибо препятствий для циркуляции крови.
Если же процесс прогрессирует, то идет инфильтрация и отложение липопротеидов, преимущественно ЛПОНП и ЛПНП в интиме артерий с последующим увеличением количества волокнистых структур межклеточного матрикса и пролиферацией клеточных элементов. В интиме возрастает количество макрофагов, которые начинают усиленно пог
лощать липопротеиды, поступающие из кровяного русла в стенку сосудов. Липопротеиды, поглощенные макрофагами, поступают в их лизосомы и там утилизируются. Но в макрофагах нет ферментных механизмов, способных расщеплять холестерол. Избыточный холестерол в клетках подвергается этерификации и откладывается в вакуолях. Эти вакуоли постепенно накапливаются в цитоплазме макрофагов, придавая цитозолю клеток ячеистый вид формируются так называемые «пенистые» клетки наличие которых в интиме артерий является характерным признаком развивающегося атеросклероза. Аналогичный процесс может, повидимому, идти и в гладкомышечных клетках, которые при развитии атеросклероза мигрируют из медии в интиму артерий, хотя с этим положением согласны не все исследователи.
Пенистые клетки гибнут, накопленный холестерол оказывается в межклеточном веществе интимы, представляя собой инородный материал. Вокруг него происходит образование соединительнотканной фиброзной капсулы, как вокруг любого чужеродного материала, попавшего в ткань. Таким путем формируется атеросклеротическая бляшка характернейший элемент атеросклеротически измененных стенок сосудов. Эта бляшка выступает в просвет сосуда, нарушая гемодинамику, бляшка может даже полностью закрывать просвет сосуда. Кроме того, изменяется моторика атеросклеротически измененных сосудов они приобретают тенденцию к спазмам, что также приводит к нарушению кровотока. Наконец, бляшки могут изъязвляться, а затем на их месте образуется рубец, деформирующий сосуд. В участках сосудистого русла с нарушенной гемодинамикой создаются условия для образования тромбов, последствиями чего и являются инфаркты и пр.
Несомненно, что в развитии атеросклеротического процесса играют роль нарушения эндотелиального слоя в крупных сосудах, в особенности ведущие к увеличению его проницаемости и возрастанию потока жидкости, а в месте с ним и потока липопротеидов, через стенку сосуда. Такие изменения наблюдаются, например, при курении или при гипертонии. Тем не менее, нарушениям липидного и в частности холестеринового обмена отводится ведущая роль в развитии атеросклероза.
В первую очередь развитию атеросклероза способствует гиперхолестеринемия.Так, по данным американских ученых у людей с содержанием холестерола в крови выше 6,7 мМ/л ( >260 мг/дл ) ишемическая болезнь сердца стенокардия и инфаркт миокарда развивается в 4 раза чаще, чем у людей с содержанием холестерола в плазме ниже 5,2 мМ/л ( 14 / 49 14 15 16 17 18 19 20 21 22 > Следующая > >>
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом