
Облегченная диффузия глюкозы из крови в ткани осуществляется при участии
Облегченную диффузию также называют диффузией с переносчиком, поскольку вещество транспортируется через мембрану с помощью специфического белка-переносчика. Таким образом, переносчик облегчает диффузию вещества на противоположную сторону мембраны.
Облегченную диффузию отличают от простой диффузии по следующей важной особенности: величина простой диффузии через открытый канал повышается пропорционально концентрации диффундирующего вещества, а при облегченной диффузии по мере повышения концентрации диффундирующего вещества скорость диффузии достигает максимума, который называют Vmax. Это различие между простой и облегченной диффузией показано на рис. 4-6. Видно, что при повышении концентрации диффундирующего вещества величина простой диффузии пропорционально возрастает, а при облегченной диффузии величина диффузии не может быть выше уровня Vmax.
Что ограничивает скорость облегченной диффузии? На этом рисунке показан белок-переносчик с порой, внешняя часть которой достаточно велика для транспорта специфической молекулы. Также показан связывающий рецептор на внутренней стороне белка-переносчика. Транспортируемая молекула входит в пору и связывается с рецептором. Затем в течение доли секунды происходит конформационное или химическое изменение в белке-переносчике, что приводит к открытию поры на противоположной стороне мембраны.
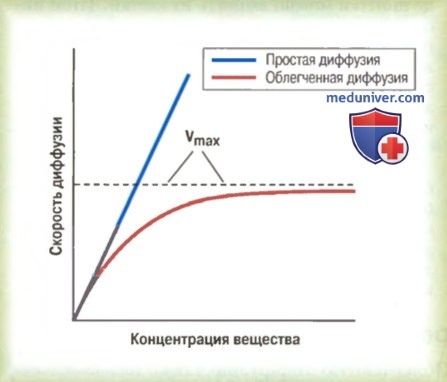
Влияние концентрации вещества на скорость диффузии через мембрану при простой и облегченной диффузии. Видно, что облегченная диффузия приближается к максимальной скорости, называемой Vmax.
Поскольку сила связи с рецептором слабая, тепловое движение прикрепленной молекулы позволяет ей оторваться от рецептора и выделиться с противоположной стороны мембраны. Скорость транспорта молекул посредством этого механизма никогда не бывает больше скорости, с которой происходят изменения молекулы белка-переносчика при переходе ее из одного состояния в другое. Следует особо отметить, что этот механизм позволяет транспортируемой молекуле двигаться, т.е. диффундировать через мембрану в любом направлении.
К наиболее важным веществам, проходящим через клеточную мембрану посредством облегченной диффузии, относят глюкозу и большинство аминокислот. Молекула переносчика для глюкозы обнаружена, и ее молекулярная масса около 45000. Она может также транспортировать некоторые другие моносахариды, структура которых подобна структуре глюкозы, включая галактозу. Кроме того, скорость облегченной диффузии глюкозы в 10-20 раз может повысить инсулин.
В настоящее время очевидно, что через клеточную мембрану могут диффундировать многие вещества. Обычно важна общая величина диффузии вещества в желаемом направлении, которая определяется рядом факторов.
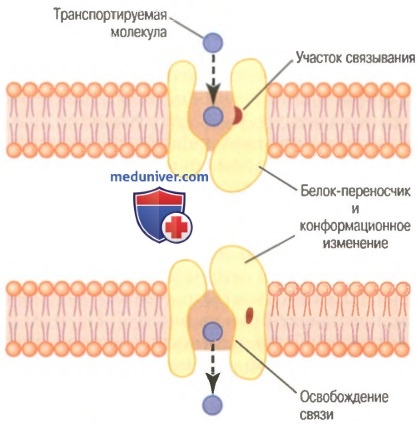
Гипотетический механизм облегченной диффузии.
Влияние разницы концентрации на величину «чистой» диффузии через мембрану. Скорость диффузии вещества внутрь пропорциональна концентрации молекул снаружи, поскольку эта концентрация определяет, как много молекул сталкивается с внешней стороной мембраны каждую секунду. Наоборот, скорость, с которой молекулы диффундируют наружу, пропорциональна их концентрации внутри мембраны. Следовательно, величина «чистой» диффузии в клетку пропорциональна разности концентраций снаружи и внутри, или
«Чистая» диффузия
(С0 — Q), где С0 — концентрация снаружи, a Q — концентрация внутри.
— Вернуться в оглавление раздела «Физиология человека.»
Облегченная диффузия
Для более крупных полярных молекул (глюкоза, аминокислоты), а также для ионов липидный бислой практически непроницаем, так как его внутренняя часть гидрофобна. Такие вещества переносятся через мембрану также по градиенту концентрации, но с участием мембранных белков.
2а. Перенос с участием ионных каналов. Трансмембранный перенос ряда ионов (Са 2+ , Na + , K + , C1 − ) происходит через ионные каналы — белковые структуры, пронизывающие мембрану. Они образуют трансмембранный гидрофильный (заполненный водой) канал. Избирательность каналов к ионам определяется наличием в белках канала специфического центра связывания иона.Каналы могут быть или закрыты, или открыты. Сигналом для изменения состояния канала может быть гормон или иная сигнальная молекула.
Читайте также: Ткань бортовка клеевая что это
2б. Перенос с помощью трансмембранных белков-переносчиков (транслоказ).Для каждого вещества или группы сходных веществ имеется свой переносчик. Переносимое вещество присоединяется к транслоказе, в результате чего изменяется ее конформация, в мембране открывается канал, и вещество освобождается с другой стороны мембраны. Поскольку в канале нет гидрофобного препятствия, то этот механизм называют облегченной диффузией. Пример — облегчённая диффузия (унипорт) глюкозы в эритроциты с помощью ГЛЮТ-1.
| Рис. 16. а — виды переноса, б-пассивный и активный транспорт. 1 — пассивная диффузия, 2 — диффузия с помощью канала, 3 — диффузия с помощью переносчика, 4 -активный транспорт, 5 — вторично-активный транспорт. |
Активный транспортвеществ протекает против концентрационного градиента и связан с расходованием энергии. Таким способом происходит перенос многих минеральных ионов из межклеточной жидкости в клетку или в обратном направлении, перенос аминокислот из просвета кишечника в клетки кишечника, перенос глюкозы из первичной мочи через клетки канальцев почки в кровь. Основным источником энергии для активного транспорта является АТФ. Поэтому, как правило, эти системы представляют собой АТФазы.
Антипорт осуществляет, например, натрий–калиевая АТФаза. Она переносит в клетку ионы калия, а из клетки — ионы натрия.
АТФ-аза присоединяет с внутренней стороны мембраны три иона Na + . Эти ионы изменяют конформацию активного центра АТФазы, и она гидролизует одну молекулу АТФ и присоединяет к себе фосфат. Выделившаяся энергия расходуется на изменение конформации АТФазы, после чего три иона натрия оказываются на внешней стороне мембраны, а фосфат замещается на 2 иона K + из внешней среды. Затем конформация переносчика изменяется на первоначальную, и ионы K + оказываются на внутренней стороне мембраны. Здесь ионы K + отщепляются, и переносчик вновь готов к работе.
Работа Na + ,K + -ATФазы создает не только разность концентраций, но и разность зарядов. На внешней стороне мембраны создается положительный заряд, на внутренней — отрицательный.
Вторично-активный транспорт.Градиент одного вещества используется для транспорта другого. Переносчик в этом случае имеет специфические центры связывания для обоих веществ. Вещество транспортируется противградиента своей концентрации путем симпорта или антипорта. Симпорт и антипорт могут происходить за счет энергии градиента концентрации ионов Na + , создаваемого Na + ,K + -ATФазой. Таким способом происходит, например, всасывание аминокислот из кишечника и глюкозы из первичной мочи икишечника.
Пример вторично-активного симпорта – транспорт глюкозы и ионов натрия; вторично-активного антипорта – Са, Nа-активный транспорт.
Для переноса углеводов, аминокислот и других метаболитов вторично-активный транспорт имеет, по-видимому, наибольшее значение по сравнению с другими механизмами.
Структура и функции мембран нарушаются при ряде заболеваний.
Контрольные вопросы
1. Перечислите основные мембранные структуры клетки.
2. Каков качественный и количественный состав мембран?
3. Какие липиды входят в состав биологических мембран? Каковы их свойства и функции?
4. В чем различие между периферическими и интегральными белками биологических мембран?
5. Какие факторы могут вызвать изменение структуры и проницаемости мембраны?
6. Перечислите виды транспорта веществ через мембраны. Какие из них требуют энергетических затрат?
7. Какую роль играют АТФ-азы в функционировании биологических мембран?
Облегченная диффузия глюкозы из крови в ткани осуществляется при участии
• Для того чтобы преодолеть гематоэнцефалический барьер, глюкоза транспортируется в астроциты через эндотелиальные клетки небольших кровеносных сосудов
Читайте также: Как удалить желтые пятна с цветной ткани
• Белки переносят глюкозу за счет унипорта, транспортируя ее в направлении градиента концентрации
• Белки, переносящие глюкозу, претерпевают конформационные изменения, которые приводят к реориентации их субстратных сайтов в мембране клетки
В клетках эукариот основным источником энергии является глюкоза, и многие клетки нуждаются в постоянном ее поступлении, поскольку у них глюкоза служит основным источником энергии для синтеза АТФ. Глюкоза представляет собой полярную молекулу, которая способна к гидратации, а клеточные мембраны не пропускают такие небольшие полярные метаболиты, как сахара. Поэтому для транспорта глюкозы в клетку требуется участие специфических мембранных белков. Транспорт глюкозы через плазматическую мембрану происходит с участием продуктов двух семейств генов.
Переносчики глюкозы (GLUTs) осуществляют независимый перенос (унипорт), при котором через мембрану происходит облегченный транспорт глюкозы. В противоположность GLUT-белкам, котранспортеры Na+/глюкоза расходуют энергию трансмембранного градиента Na+ на транспорт глюкозы. В настоящем разделе мы рассмотрим GLUT-белки.
![]()
Глюкоза селективно транспортируется через гематоэнцефалический барьер посредством изоформы 1 (GLUT-1) белка-переносчика.
Перенос глюкозы из крови в головной мозг и в другие отделы ЦНС представляет собой многоступенчатый процесс,
в котором участвуют различные типы клеток.
Семейство GLUT-белков является частью более обширного суперсемейства белков, облегчающих основные транспортные процессы (MFS) и широко представленных в клетках всех организмов. GLUT-белки представляют собой интегральные белки мембран клеток эукариот. Изоформы переносчиков GLUT различаются по своим кинетическим характеристикам, специфичностью по отношению к переносимым сахарам, тканевой локализацией и механизмами регуляции. Некоторые GLUT-белки, наряду с глюкозой, транспортируют и другие метаболиты, например галактозу, воду и анальгетики из группы гликопептидов.
GLUT-белки осуществляют унипорт, при котором вещества проходят через мембрану в направлении градиента концентрации. Таким образом, в зависимости от концентрации, GLUT-белки переносят метаболиты в клетку или в противоположном направлении. Поступление в клетку глюкозы, происходящее при участии GLUT-белков, часто определяет жизнеспособность клеток, которые характеризуются высоким уровнем потребления энергии.
Питательные метаболиты, такие как сахара, через кровеносные сосуды транспортируются в органы. Эндотелиальные клетки, выстилающие стенки небольших сосудов, контролируют процессы обмена питательных веществ. В этих эндотелиальных клетках, особенно расположенных в области гематоэнцефалического барьера, содержится много GLUT-белков. Надлежащее функционирование головного мозга сильно зависит от глюкозы, и его клетки особенно чувствительны к снижению ее содержания.
Транспорт глюкозы в нервные клетки происходит через капилляры мозга в несколько этапов и с участием изоформы GLUT-1. Эта изоформа экспрессируется в мембране клеток эндотелия, находящихся на границе между кровью и межклеточным пространством, а также в плазматической мембране астроцитов, функция которых важна в гематоэнцефалическом барьере. Расположенные в этих местах белки GLUT-1 транспортируют глюкозу из крови в эндотелиальные клетки, и оттуда в астроциты. В них глюкоза превращается в другие источники энергии, которые транспортируются в нейроны.
В различных тканях содержатся разные изоформы GLUT. Например, в клетках мышечной и жировой ткани транспорт глюкозы осуществляется с участием GLUT-4. В процессе приема пищи или после еды в клетки этих тканей под действием инсулина увеличивается поступление глюкозы. При этом происходит регулируемый транспорт изоформы GLUT-4 к клеточной поверхности. Эта изоформа также называется инсулин-зависимый переносчик. Белок GLUT-4 локализован во внутриклеточных везикулах, которые сливаются с плазматической мембраной. Этим обеспечивается доставка переносчика GLUT-4 к плазматической мембране и увеличивается емкость транспортного процесса.
Читайте также: Костная ткань спирального органа составляет
![]()
Предполагаемое строение GLUT-переносчика, состоящего из 12 трансмембранных сегментов с внутриклеточными С- и N-концевыми участками.
Внутриклеточные петли содержат сайты фосфорилирования и связывания субстратов.
Показано, что градиент глюкозы по обеим сторонам мембраны может возникать в любом направлении, в зависимости от типа клеток и их метаболизма.
Градиент определяет направление транспорта.
Связывание инсулина со своим поверхностным рецептором запускает каскад внутриклеточных процессов, приводящих к быстрому слиянию этих везикул с мембраной. Это, в свою очередь, приводит к быстрому увеличению транспорта глюкозы в клетку с участием GLUT-4. При диабете типа II не происходит транспорт глюкозы из крови в мышцы и жировую ткань, очевидно, из-за нарушения позиционирования GLUT-4 на плазматической мембране. Более того, GLUT-2 способен экспортировать глюкозу из клеток тех органов, в которых она образуется, например из клеток печени.
По строению переносчик GLUT напоминает другие члены суперсемейства MFS. Предполагается, что он состоит из 12 трансмембранных а-спиралей, имеющих внутриклеточные N- и С-концевые участки и петли. Последние содержат сайт связывания субстрата и сайты фосфорилирования.
Модель структуры GLUT-1 предложена на основании результатов по сайт-направленному мутагенезу и по измерению транспорта глюкозы мутантным белком. Для построения модели также использовались данные по кристаллической структуре бактериальной лактопермеазы, относящейся к подсемейству переносчиков олигосахари-дов/Н+, входящих в MFS.
Согласно этой модели, при соответствующей ориентации трансмембранных спиралей создается полость поры, через которую транспортируется глюкоза, а также обеспечивается образование водородных связей между GLUT-белком и глюкозой. Результаты кинетического анализа транспорта глюкозы в эритроцитах позволяют предполагать существование специального механизма, который обеспечивает белку GLUT возможность принимать одну из двух основных конформаций. Этот механизм аналогичен постулируемому для бактериальной лактопермеазы. Когда белок находится в одной конформации, сайт связывания глюкозы обращен в сторону внеклеточного пространства. В другой конформации он обращен в сторону цитозоля. При связывании глюкозы с любой стороны происходят конформационные изменения, которые приводят к реориентации сайтов связывания глюкозы к противоположной стороне мембраны и к ее высвобождению.
Таким образом, хотя переносчики GLUT являются унипортерами, а бактериальная лактопермеаза представляет собой симпортер, предполагается, что они функционируют по одному механизму.
Мутации в гене GLUT-1 вызывают значительные дефекты развития. У детей потребность головного мозга в глюкозе в 3-4 раза выше, чем у взрослых, и в мозг поступает до 80% всей глюкозы. Мутации в гене GLUT-1 человека служат причиной проявления редкого синдрома недостаточности GLUT-1, который характеризуется развитием припадков и замедлением развития. Считается, что это связано с нарушением транспорта глюкозы в головной мозг. Эмбрионы мышей, дефектных по гену GLUT-1, отстают в росте, и у них обнаруживаются различные пороки развития. Аналогичные дефекты возникают у мышиных эмбрионов, которые развиваются в организме диабетических самок. При этом избыток глюкозы в крови подавляет экспрессию GLUT-1 в органах эмбриона.
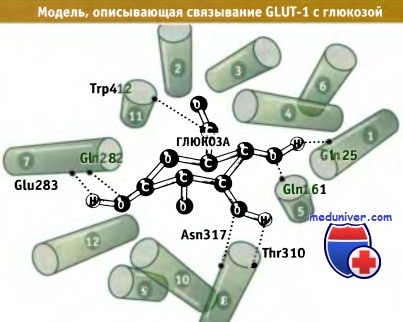
Предполагаемая ориентация трансмембранных а-спиралей переносчика глюкозы GLUT-1.
Показаны остатки, участвующие в связывании глюкозы (размер молекулы глюкозы представлен без учета масштаба).
Вид на спирали со стороны внутренней части плазматической мембраны.
Эта модель построена с использованием гомологичной структуры лактопермеазы E.coli в качестве матрицы.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом