
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
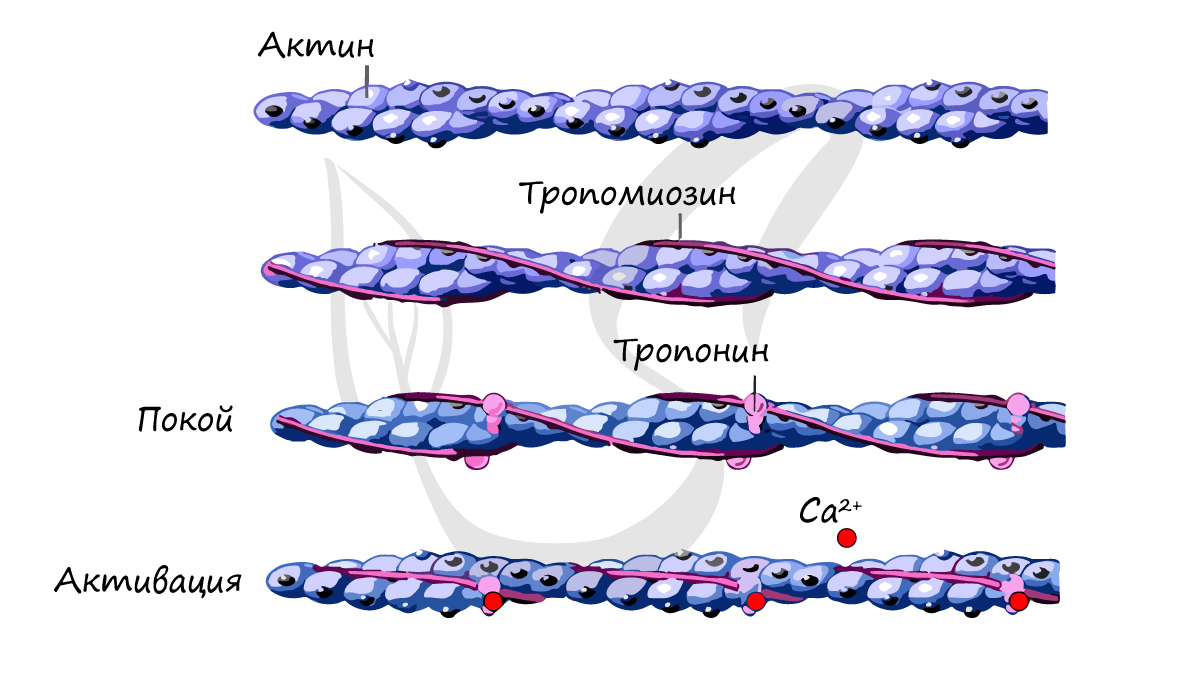
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.
Читайте также: Техника исполнения мягкой игрушки из ткани

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Глава 9. Мышечные ткани
Мышечными тканями (textus muscularis) называют ткани, различные по строению и происхождению, но сходные по способности к сокращениям. Они обеспечивают перемещения в пространстве организма в целом, его частей и движение органов внутри организма (сердце, язык, кишечник и др.).
Свойством сокращаться с изменением формы обладают клетки многих тканей, но в мышечных тканях эта способность становится главной функцией.
9.1. Общая морфофункциональная характеристика и классификация
Основные морфологические признаки элементов мышечных тканей — удлиненная форма, наличие продольно расположенных миофибрилл и миофиламентов — специальных органелл, обеспечивающих сократимость, расположение митохондрий рядом с сократительными элементами, наличие включений гликогена, липидов и миоглобина.
Специальные сократительные органеллы — миофиламенты или миофи-бриллы, обеспечивают сокращение, которое возникает при взаимодействии в них двух основных фибриллярных белков — актина и миозина, при обязательном участии ионов кальция. Митохондрии обеспечивают эти процессы энергией. Запас источников энергии образуют гликоген и липиды. Миоглобин — белок, обеспечивающий связывание кислорода и создание его запаса на момент сокращения мышцы, когда сдавливаются кровеносные сосуды (поступление кислорода при этом резко снижается).
Классификация. В основу классификации мышечных тканей положены два принципа — морфофункциональный и гистогенетический. В соответствии с морфофункциональным принципом, в зависимости от структуры органелл сокращения, мышечные ткани подразделяют на две подгруппы.
Первая подгруппа — поперечнополосатые (исчерченные) мышечные ткани (textus muscularis striatus). В цитоплазме их элементов миозиновые филамен-
ты постоянно полимеризованы, образуют с актиновыми нитями постоянно существующие миофибриллы. Последние организованы в характерные комплексы — саркомеры. В соседних миофибриллах структурные субъединицы саркомеров расположены на одном уровне и создают поперечную исчер-ченность.
Вторая подгруппа — гладкие (неисчерченные) мышечные ткани (textus muscularis nonstriatus). Эти ткани характеризуются тем, что вне сокращения миозиновые филаменты деполимеризованы. В присутствии ионов кальция они полимеризуются и вступают во взаимодействие с филаментами актина. Образующиеся при этом миофибриллы не имеют поперечной исчерченности: при специальных окрасках они представлены равномерно окрашенными по всей длине (гладкими) нитями.
Читайте также: Ткань жаккард для штор характеристики
В соответствии с гистогенетическим принципом в зависимости от источников развития (эмбриональных зачатков) мышечные ткани и мышечные элементы подразделяются на: соматические (миотомные), целомические (из миоэпикардиальной пластинки висцерального листка спланхнотома), мезенхимные (из десмального зачатка в составе мезенхимы), нейральные (из нервной трубки), эпидермальные (из кожной эктодермы и из прехор-дальной пластинки).
9.2. ПОПЕРЕЧНОПОЛОСАТЫЕ МЫШЕЧНЫЕ ТКАНИ
Имеется две основные разновидности поперечнополосатых (исчерченных) тканей — скелетная (миотомная) и сердечная (целомическая).
9.2.1. Скелетная мышечная ткань
Гистогенез. Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани (textus muscularis striatus sceletalis) являются стволовые клетки миотомов — промиобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтон-ных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты — мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл — миофибрилл (рис. 9.1). В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндо-

Рис. 9.1. Гистогенез скелетной мышечной ткани (по А. А. Клишову):
а — промиобласты; б — миосимпласт; в — мышечная трубочка; г — зрелое мышечное
волокно. 1 — миосателлитоцит; 2 — ядро миосимпласта; 3 — миофибриллы
плазматическая сеть в значительной степени редуцируется. Такие дефинитивные структуры называют миосимпластами.
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты. Эти клетки располагаются на поверхности миосимпластов. Миосателлитоциты, размножаясь, сливаются с миосимпластами, участвуя таким образом в создании оптимального ядерно-

Рис. 9.2. Строение поперечнополосатой мышечной ткани (микрофотография):
1 — мышечные волокна; 2 — сарколемма; 3 — саркоплазма и миофибриллы; 4 — ядра
миосимпласта. Окраска — железный гематоксилин
саркоплазменного отношения, необходимого для синтеза специфических белков симпласта.
Строение. Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосател-литоцитов, покрытых общей базальной мембраной (рис. 9.2-9.4). Длина всего волокна может измеряться сантиметрами при толщине 50-100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Строение миосимпласта. Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под плазмолеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч (см. рис. 9.2). У полюсов ядер располагаются органеллы общего значения — комплекс Гольджи и небольшие фрагменты агранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно (см. рис. 9.3).
Саркомер — структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски с неодинаковым лучепреломлением (анизотропные А-диски и изотропные I-диски). Каждая миофибрил-ла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети — саркоплазмати-ческой сети. Соседние саркомеры имеют общую пограничную структуру — Z-линию (рис. 9.5). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы актиновых филаментов. От соседних Z-линий акти-новые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина

Рис. 9.3. Схема ультрамикроскопического строения миосимпласта (по Р. В. Крстичу, с изменениями) (а): 1 — саркомер; 2 — анизотропный диск (полоса А); 2а — изотропный диск (полоса I); 3 — линия М (мезофрагма) в середине анизотропного диска; 4 — линия Z (телофрагма) в середине изотропного диска; 5 — митохондрии; 6 — сар-коплазматическая сеть; 6а — конечная цистерна; 7 — поперечная трубочка (Т-трубочка); 8 — триада; 9 — сарколемма; б — схема пространственного расположения митохондрий в симпласте. Верхняя и нижняя плоскости рисунка ограничиваютанизотропныйдисксарко-мера (по Л. Е. Бакеевой, В. П. Скулачеву, Ю. С. Ченцову); в — эндомизий. Сканирующая электронная микрофотография, увеличение 2600 (препарат Ю. А. Хорошкова): 1 — мышечные волокна; 2 — коллагеновые фибриллы
фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомиозина. Она образует в сечении М-линию. В узлах этой М-линии закреплены концы мио-зиновых филаментов. Другие их концы направляются в сторону Z-линий и

Рис. 9.4. Поверхностный участок миосимпласта и миосателлитоцит. Электронная микрофотография, увеличение 10 000 (препарат В. Л. Горячкиной и С. Л. Кузнецова): 1 — базальная мембрана; 2 — плазмолемма; 3 — ядро миосимпласта; 4 — ядро миоса-теллитоцита; 5 — миофибриллы; 6 — канальцы агранулярной эндоплазматической (саркоплазматической) сети; 7 — митохондрии; 8 — гликоген

1 — линия Z; 2 — линия М; 3 — филаменты актина; 4 — филаменты миозина; 5 — фибриллярные молекулы титина (по Б. Албертс, Д. Брей, Дж. Льюис и др., с изменениями)

Рис. 9.6. Конформационные изменения, влекущие за собой взаимное смещение филаментов актина и миозина:
а-в — последовательные изменения пространственных отношений. 1 — актин; 2 — головка молекулы миозина (по Б. Албертс, Д. Брей, Дж. Льюис и соавт., с изменениями)
располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
Молекулы миозина имеют длинный хвост и на одном из его концов две головки. При повышении концентрации ионов кальция в области присоединения головок (шарнирный участок) молекула изменяет свою конфигурацию (рис. 9.6). При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Читайте также: Заводы для производства тканей
Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в ее кортикальном слое, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна.
Источником ионов кальция служат цистерны агранулярной эндоплазма-тической сети. Они вытянуты вдоль миофибрилл около каждого саркоме-ра и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или латеральные (L) цистерны.
С поверхности в глубину миосимпласта плазмолемма образует длинные трубочки, идущие поперечно (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда миосимпласт получает сигнал о начале сокращения, он перемещается по плазмолемме в виде потенциала действия и распространяется на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в канальцах сети и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет превращения АТФ в АДФ. Роль АТФ-азы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами.
Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма. Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается и участвует в биохимических реакциях.
Миосателлитоциты. Эти малодифференцированные клетки являются источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются (см. рис. 9.1, 9.4). Миосателлитоциты одноядерны, их темные ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром).
Типы мышечных волокон. Разные мышцы (как органы) функционируют в неодинаковых биомеханических условиях. Поэтому и мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Активность ферментов в них различна, и представлены они в различных изомерных формах. Различно в них и содержание дыхательных ферментов — гликолитических и окислительных.

Рис. 9.7. Активность сукцинатдегидрогеназы в мышечных волокнах разного типа (препарат В. Ф. Четвергова, обработка по Нахласу и соавт.): 1 — высокая; 2 — низкая; 3 — средняя
По соотношению миофибрилл, митохондрий и миоглобина различают белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют на быстрые, медленные и промежуточные, что определяется молекулярной организацией миозина. Среди его изоформ существуют две основные — «быстрая» и «медленная». При постановке гистохимических реакций их идентифицируют по АТФ-азной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах преобладают гликолитические процессы, они богаты гликогеном, в них меньше миоглобина, поэтому их называют белыми. В медленных волокнах, напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными.
Наряду с белыми и красными существуют и промежуточные волокна. В составе большинства скелетных мышц волокна разного гистохимического типа располагаются мозаично (рис. 9.7).
Свойства мышечных волокон меняются при изменении нагрузок — спортивных, профессиональных, а также в экстремальных условиях (невесомость). При возврате к обычной деятельности такие изменения обратимы. При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц.
Регенерация. Ядра миосимпластов делиться не могут, так как в саркоплазме отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки сливаются с миосимпластами. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагмен-
ты фагоцитируются макрофагами. Восстановление тканей осуществляется за счет двух механизмов: компенсаторной гипертрофии самого симпласта и пролиферации миосателлитоцитов. В симпласте активизируются гранулярная эндоплазматическая сеть и комплекс Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмо-леммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку. Миосателлитоциты, сохранившиеся рядом с повреждением, делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют новые миотубы, которые развиваются в мышечных волокна.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом