
Основные продукты метаболизма глюкозы в ткани печени
Глюкоза, наряду с жирными кислотами и кетоновыми тепами, является важнейшим источников энергии. Уровень глюкозы в крови поддерживается постоянным 4-6 мМ (0,8-1,0 г/л) благодаря тонкой регуляции процессов ее поступления и потребления. Глюкоза поступает из кишечника (за счет переваривания пищи), печени и почек. При этом печень выполняет функцию «глюкостата»: в фазе резорбции глюкоза поступает в печень из крови и накапливается в виде гликогена. При дефиците глюкозы ( фаза пострезорбции , голодание) печень, напротив, поставляет глюкозу, которая образуется за счет процессов гликогенолиза и глюконеогенеза (см. с. 300).
Печень обладает свойством синтезировать глюкозу из других сахаров, например фруктозы и галактозы, или из других продуктов промежуточного метаболизма. Превращение лактата в глюкозу в цикле Кори (см. с. 330) и аланина в глюкозу в цикле аланина (см. с. 330) играет особую роль в обеспечении эритроцитов и мышечных клеток.
Необходимыми условиями активного углеводного обмена в печени является обратимый транспорт сахаров через плазматическую мембрану гепатоцитов (при отсутствии контроля инсулином) и наличие фермента глюкозо-6-фосфатазы, высвобождающего глюкозу из глюкозо-6-фосфата.
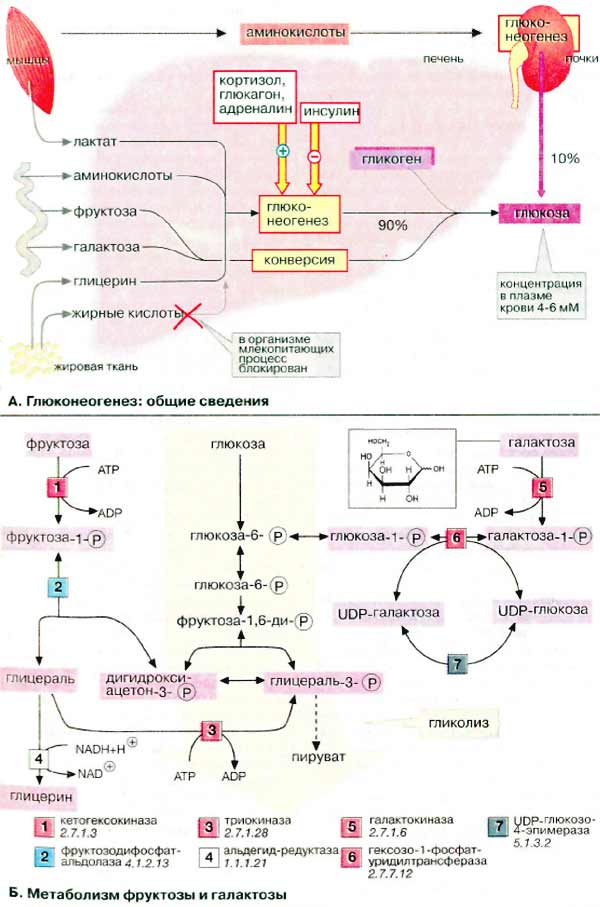
А. Глюконеогенез: общие сведения
Синтез глюкозы de novo (до 250 г в сутки) происходит в основном в печени. Процесс глюконеогенеза может идти и в почках, однако из-за небольших размеров почек их вклад в синтез глюкозы составляет всего 10%.
Глюконеогенез контролируется гормонами. Кортизол, глюкагон и адреналин стимулируют этот процесс, а инсулин, напротив, подавляет.
При глюконеогенезе в печени наиболее важными субстратами являются лактат, поступающий из мышечной ткани и эритроцитов, аминокислоты из желудочно-кишечного тракта (глюкогенные аминокислоты) и мышц (аланин), а также глицерин из жировых тканей. В почках в качестве субстрата служат главным образом аминокислоты (см. с. 320).
Жирные кислоты и другие источники ацетил-КоА не могут использоваться в организме млекопитающих для биосинтеза глюкозы, поскольку ацетил-КоА, образующийся при β-окислении в цитратном цикле (см. с. 140), полностью окисляется до СО 2 , в то время как в глюконеогенезе исходным продуктом является оксалоцетат.
Б. Метаболизм фруктозы и галактозы
Метаболизм фруктозы осуществляется превращением ее в глюкозу (на схеме слева). Вначале фруктоза фосфорилируется при участии фермента кетогексокиназы (фруктокиназы) [ 1 ] с образованием фруктозо-1-фосфата, который далее расщепляется альдолазой до глицеральдегида (глицераля) и дигидроксиацетон-3-фосфата [ 2 ]. Последний уже является промежуточным продуктом гликолиза (в центре схемы), а глицераль фосфорилируется в присутствии триокиназы, образуя глицераль-3-фосфат [ 3 ].
Затем глицеральдегид частично восстанавливается до глицерина [ 4 ] или окисляется до глицерата. После фосфорилирования оба соединения вновь включаются в гликолиз (на схеме не приведено). При восстановлении глицеральдегида [ 4] расходуется НАДН (NADH). Поскольку при конверсии этанола лимитирующим фактором является низкое соотношение концентраций НАД + /НАДН (NAD + /NADH). Этот процесс ускоряется в присутствии фруктозы (см. с. 312).
Кроме того, в печени реализуется полиольный путь трансформации фруктозы в глюкозу (на схеме не приведен): фруктоза за счет восстановления С-2 превращается в сорбит, а при последующем дегидрировании С-1 — в глюкозу.
Метаболизм галактозы также начинается с фосфорилирования с образованием галактозо-1-фосфата [ 5 ] (на схеме справа). Далее следует эпимеризация С-4 с образованием производного глюкозы. Биосинтез УДФ-глюкозы (UDP-глюкозы), промежуточного продукта обмена глюкозы, осуществляется обходным путем — через УДФ-галактозу (UDP-галактозу) и последующую эпимеризацию [ 6, 7 ]. По такому же пути идет биосинтез самой галактозы, поскольку все реакции за исключением [ 5 ] обратимы.
48. Особенности обмена глюкозы в разных органах и клетках: эритроциты, мозг, мышцы, жировая ткань, печень.
Метаболизм глюкозы в эритроцитах. Эритроциты лишены митохондрий, поэтому в качестве энергетического материала они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроциты путём облегчённой диффузии с помощью ГЛЮТ-2. Около 90% поступающей глюкозы используется в анаэробном гликолизе, а остальные 10% — в пентозофосфатном пути. Конечный продукт анаэробного гликолиза лактат выходит в плазму крови и используется в других клетках, прежде всего гепатоцитах. АТФ, образующийся в анаэробном гликолизе, обеспечивает работу Nа + , К+-АТФ-азы и поддержание самого гликолиза, требующего затраты АТФ в гексокиназной и фосфофруктокиназной реакциях. Важная особенность анаэробного гликолиза в эритроцитах по сравнению с другими клетками — присутствие в них фермента бисфосфоглицератмутазы. Бисфосфоглицератмутаза катализирует образование 2,3-бисфосфоглицерата из 1,3-бисфосфоглицерата. Образующийся только в эритроцитах 2,3-бисфосфоглицерат служит важным аллостерическим регулятором связывания кислорода гемоглобином. Глюкоза в эритроцитах используется и в пентозофосфатном пути, окислительный этап которого обеспечивает образование кофермента NADPH, необходимого для восстановления глу-татиона.
Читайте также: Ткань пальтовый в уфе
Метаболизм глюкозы в печени.Основная рольпеченив углеводном обмене заключается в обеспечении постоянстваконцентрацииглюкозывкрови. Это достигается регуляцией междусинтезом и распадом гликогена, депонируемого впечени. Впеченисинтез гликогенаи его регуляция в основном аналогичны тем процессам, которые протекают в других органах итканях, в частности вмышечной ткани.Синтез гликогенаизглюкозыобеспечивает в норме временный резервуглеводов, необходимый для поддержанияконцентрацииглюкозывкровив тех случаях, если ее содержание значительно уменьшается (например, у человека это происходит при недостаточном поступленииуглеводовс пищей или в период ночного «голодания»). Необходимо подчеркнуть важную рольферментаглюкокиназы в процессе утилизацииглюкозыпеченью. Глюкокиназа, подобногексокиназе, катализируетфосфорилированиеглюкозыс образованием глюкозо-6-фосфата, при этомактивностьглюкокиназы впеченипочти в 10 раз превышаетактивностьгексокиназы. Важное различие между этими двумяферментамизаключается в том, что глюкокиназа в противоположностьгексокиназеимеет высокое значение КМ дляглюкозыи не ингибируется глюкозо-6-фосфатом. Считают, что основная рольпечени– расщеплениеглюкозы– сводится прежде всего к запасанию метаболитов-предшественников, необходимых длябиосинтеза жирных кислотиглицерина, и в меньшей степени кокислениюее до СО2 и Н2О. Синтезированные впеченитриглицериды в норме выделяются вкровьв составелипопротеинови транспортируются вжировую тканьдля более «постоянного» хранения. Вреакцияхпентозофосфатного пути впечениобразуется НАДФН, используемый для восстановительныхреакцийв процессах синтезажирных кислот,холестеринаи другихстероидов. Кроме того, при этом образуются пентозофосфаты, необходимые для синтезануклеиновых кислот. Наряду с утилизациейглюкозывпеченипроисходит и ее образование. Непосредственным источникомглюкозывпеченислужитгликоген.Распад гликогенавпеченипроисходит в основном фосфоролитическим путем. В регуляции скоростигликогенолизавпеченибольшое значение имеет система циклическихнуклеотидов. Кроме того,глюкозавпечениобразуется также в процессеглюконеогенеза.
Метаболизм глюкозы в мозгу.Основнымсубстратомдыханиямозговойтканиявляетсяглюкоза. В 1 мин 100 гтканимозга потребляют в среднем 5 мгглюкозы. Подсчитано, что более 90% утилизируемойглюкозывтканимозга окисляется до СО2и Н2О при участиицикла трикарбоновых кислот. В физиологических условиях роль пентозофосфатного путиокисленияглюкозыв мозговойтканиневелика, однако этот путьокисленияглюкозыприсущ всемклеткамголовного мозга. Образующаяся в процессепентозофосфатного циклавосстановленная форма НАДФ (НАДФН) используется для синтезажирных кислотистероидов. Междуглюкозойигликогеноммозговойтканиимеется тесная связь, выражающаяся в том, что при недостаточном поступленииглюкозыизкровигликогенголовного мозга является источникомглюкозы, аглюкозапри ее избытке – исходным материалом длясинтеза гликогена.Распад гликогенав мозговойтканипроисходит путем фосфоролиза с участием системыцАМФ. Однако в целом использованиегликогенав мозге по сравнению сглюкозойне играет существенной роли в энергетическом отношении, так как содержаниегликогенав головном мозге невелико. Наряду с аэробнымметаболизмом углеводовмозговаятканьспособна к довольно интенсивному анаэробномугликолизу. Значение этого явления пока недостаточно ясно, ибогликолизкак источник энергии ни в коей мере не может сравниться по эффективности с тканевымдыханиемв головном мозге.
Метаболизм глюкозы в мышцах.При работе умеренной интенсивности мышца может покрывать свои энергетические затраты за счет аэробногометаболизма. Однако при больших нагрузках, когда возможность снабжениякислородомотстает от потребности в нем, мышца вынуждена использовать гликолитический путь снабжения энергией. При интенсивной мышечной работе скорость расщеплениягликогенаилиглюкозыс образованиеммолочной кислотыувеличивается в сотни раз. Соответственно содержаниемолочной кислотывмышечной тканиможет повышаться до 1,0–1,2 г/кг и более. С токомкровизначительное количествомолочной кислотыпоступает впечень, где ресинтезируется вглюкозуигликоген(глюконеогенез) за счет энергии окислительных процессов.
Метаболизм глюкозы в жировой ткани. В адипоцитах для обеспечения реакций синтеза жира распад глюкозы идёт по двум путям: гликолиз, обеспечивающий образование глицерол-3-фосфата и ацетил-КоА, и пентозофосфатный путь, окислительные реакции которого обеспечивают образование NADPH, служащего донором водорода в реакциях синтеза жирных кислот.
Основные продукты метаболизма глюкозы в ткани печени
Недостаточность углеводного обмена в печени, нарушение ее глюкостатической функции может носить первичный наследственный и вторичный приобретенный характер. Причем приобретенные формы печеночной недостаточности в поддержании углеводного гомеостаза могут возникать как следствие прямого цитотоксического действия на паренхиму печени патогенных факторов бактериально-токсической, иммунноаллергической, онкогенной природы экзогенного или эндогенного происхождения. В ряде случаев расстройства углеводного обмена в печени возникают в результате нарушений оксигенации паренхимы печени или развития холестаза [1–5].
Читайте также: Ткань для реставрации одежды
Следует отметить важное значение нарушений гормонального баланса, в частности изменений соотношения в организме инсулина и контринсулярных гормонов в расстройствах процессов гликолиза, гликогенолиза, гликогенеза, гликонеогенеза, окисления субстратов в цикле Кребса и других реакциях.
Характер и механизмы нарушений углеводного обмена в печени при наследственных формах патологии
Наследственные нарушения метаболизма углеводов в печени именуют гликогенозами (болезни накопления) и агликогенозами (отсутствие гликогена в печени).
Следует отметить, что гликогенозы могут быть не только печеночного происхождения, но и мышечного, а также носить смешанный и даже генерализованный характер.
Первая идентификация гликогенозов и их классификация была предложена супругами Cori (1957). В последующем было установлено XI различных энзиматических дефектов метаболизма углеводов, приводящих к качественным и количественным нарушениям содержания гликогена в органах и тканях. Наиболее часто встречающимися гликогенозами являются гепаторенальный гликогеноз Iа типа (болезнь Гирке), описанный впервые von Girke в 1929 г. [2, 5].
Заболевание выявляется в раннем детском возрасте, характеризуется резким снижением содержания глюкозы в крови (иногда до 0,55–1,1 ммоль/л) и сопровождается развитием судорог, рвоты, коллапса.
Основной генетический дефект, лежащий в основе данного заболевания – это недостаточность фермента глюкозо-6-фосфатазы, наследуемая аутосомно-рецессивно. При этом в ткани печени накапливается глюкозо-6-фосфат, активирующий Д-форму гликогенсинтетазы. Последнее приводит к прогрессирующему возрастанию содержания гликогена в печени. Одновременно выявляется избыточное отложение гликогена в канальцах почек.
Снижение содержания глюкозы в крови подавляет продукцию инсулина, что приводит к активации липолиза, развитию гиперлипидемии, нарушению утилизации субстратов в цикле Кребса. Одновременно возникает накопление промежуточных продуктов распада углеводов и жиров: молочной, пировиноградной кислоты, жирных и кетокислот, а также холестерина, триглециридов, мочевой кислоты.
Достаточно быстро формируется метаболический ацидоз. Формирование гепаторенального гликогеноза нередко приводит к развитию таких осложнений, как подагра, уролитиаз.
Касаясь нарушений клеточного состава периферической крови при гликогенозе Iа типа, следует отметить развитие анемии и лейкопении, при этом в лейкоцитах резко увеличивается содержание гликогена.
Прогноз заболевания неблагоприятен вплоть до развития летальных исходов (в 50 % наблюдений) в раннем детском возрасте. Однако в других случаях возможно относительно благоприятное течение патологии и рождение здоровых детей в потомстве.
Гликогеноз Ib типа характеризуется наследственной недостаточностью транслоказы глюкозо-6-фосфатазы в эндоплазматическом ретикулюме гепатоцитов, в то время как активность фермента глюкозо-6-фосфатазы достаточно велика.
Гликогеноз III типа (болезнь Форбса) связан с недостаточностью фермента амило-1-6-глюкозидазы, является наиболее частой формой гликогеноза, характеризуется выраженной гепатомегалией гипокликемией, умеренным ацидозом и кетонурией. У детей отмечается умеренная задержка роста. Тем не менее течение этой формы патологии более благоприятное, чем при болезни Гирке.
Гликогеноз IV типа (болезнь Андерсена) – крайне редкое аутосомно-рецессивное заболевание связано с наследственным дефектом ветвящего фермента гликогена не только в печени, но в лейкоцитах, фибробластах и мышцах. Содержание гликогена в печени при указанной патологии чаще нормальное, однако структура его аномальна, сходна с амилопектином, включает редкие длинные наружные ветви.
Гипогликемический синдром не выражен, однако прогноз заболевания неблагоприятен в связи с ранним развитием цирроза печени.
Гликогеноз VIа типа (болезнь Хёрса) – редкая форма аутосомно-рецессивного наследования нарушения активности фосфорилазы печени и лейкоцитов. Заболевание характеризуется развитием гипогликемии и умеренного ацидоза. Прогноз болезни благоприятен в связи с восполнением дефекта фермента фосфорилазы за счет активации реакций глюконеогенеза.
Этиология и патогенез агликогеноза
Агликогеноз – заболевание с аутосомно-рецессивной недостаточностью УДФ-глюкозо-гликогентрансферазы (гликогенсинтетазы) характеризуется отсутствием запасов гликогена в печени, развитием гипогликемии, особенно натощак, в связи с чем возникает необходимость частого приема пищи. В ряде случаев формируется жировая инфильтрация печени, возрастает уровень кетоновых тел в крови. Одновременно снижается содержание аминокислот в крови, участвующих в глюконеогенезе.
Характер и механизмы нарушений углеводного обмена в печени при приобретенных формах патологии
Как указывалось выше, печень играет ведущую роль в поддержании постоянства содержания глюкозы в крови за счет динамического равновесия процессов гликогенеза, глюконеогенеза, гликолиза и гликогенолиза. При повреждении паренхимы печени прежде всего нарушается глюкостатическая функция. Это обусловлено тем фактом, что гликогенез является энергозависимым процессом. Удлинение цепи гликогена на одну молекулу глюкозы требует расхода одной молекулы АТФ и УДФ. В то же время развитие деструктивно-воспалительных процессов гепатоцитов, нарушение оксигенации и трофики печени при различных формах патологии инфекционной и неинфекционной природы, нарушения гормонального баланса, кислотно-основного состояния сопровождаются развитием типовой реакции митохондрий – набуханием [5–8].
Читайте также: Чем закрепить цвет ткани если она линяет
При набухании митохондрий нарушается пространственная ориентация дыхательных ферментативных ансамблей на внутренней мембране митохондрий, разобщаются процессы окислительного фосфорилирования и дыхания, возникает дефицит АТФ и, соответственно, подавление всех энергозависимых реакций, в том числе и гликогенеза. В этих случаях нагрузка глюкозой приводит к появлению длительной диабетоидной гипергликемической кривой, а вне нагрузки глюкозой или при редком приеме пищи в связи с истощением запасов гликогена возможна гипогликемия.
Пораженная печень теряет способность трансформировать в гликоген и другие субстраты в реакциях гликонеогенеза, что приводит к избыточному накоплению в крови, тканях и паренхиме печени молочной, пировиноградной, жирных кислот, кетокислот, аминокислот. Последнее в свою очередь является одним из патогенетических факторов развития метаболического ацидоза. Для печеночной недостаточности характерно также нарушение окисления глюкозы в пентозном цикле и соответственно развитие дефицита рибозофосфатов, а также НАДФН2, что приводит к недостаточности антиоксидантных систем и активации свободнорадикальной деструкции гепатоцитов. Нарушения трансформации пировиноградной кислоты в гликоген и окисления ее в цикле трикарбоновых кислот при печеночной недостаточности сопровождается активацией альтернативных механизмов ее метаболизма с образованием токсических соединений – ацетона и бутиленгликоля.
Нарушения углеводного обмена в печени при гормональном дисбалансе
Основными гормональными регуляторами углеводного обмена в печени являются инсулин и контринсулярные гормоны (глюкогон, адреналин, норадреналин, тираксин, трийодтиранин, соматотропный гормон и адренокортикотопный гормоны).
В то же время следует отметить, что гепатоциты занимают промежуточное положение между клетками высокоинсулинзависимых органов и тканей (жировой, соединительной, мышечной, кроветворной и др.) и инсулиннезависимых (центральной нервной системы, надпочечников, гонад, глаза) [2, 3, 5].
Как известно, плазматическая мембрана гепатоцита, в отличие от мио- и липоцита, свободно проницаема для глюкозы без участия инсулина. В то же время в условиях нормы под влиянием инсулина возникает активация гексокиназы (глюкокиназы) в гепатоцитах, миоцитах и липоцитах, а также усиливается синтез жирных кислот из глюкозы, активируются пентозные и гликолитические пути метаболизма глюкозы, повышается активность ферментов цикла Кребса. Активация указанных реакций обеспечивает усиление синтеза АТФ. Дефицит инсулина, как известно, может носить наследственный и приобретенный характер, быть абсолютным или относительным, иметь панкреатическое или внепанкреатическое происхождение. В настоящее время очевидна возможность развития сахарного диабета I и II типов как самостоятельных нозологических форм патологии наследственного генеза, а также формирование вторичного сахарного диабета симптоматического характера при различных эндокринопатиях, сопровождающихся гиперпродукцией контринсулярных гормонов, а также при воспалительно-деструктивных процессах в поджелудочной железе неаутоиммунного характера (при несбалансированном питании, недостаточности незаменимых аминокислот, витаминов, микроэлементов и т.д.) [6, 9, 10].
В условиях дефицита инсулина, абсолютного или относительного преобладания контринсулярных гормонов при различных эндокринопатиях подавляются процессы гликогенеза, снижается активность ферментов пентозного цикла окисления глюкозы, а также окисление субстратов в цикле Кребса. Гликонеогенез активируется лишь под влиянием гормонов АКТГ и глюкокортикоидов при стрессорных ситуациях, болезни и синдроме Иценко – Кушинга, стероидной терапии. Между тем усиление продукции контринсулярных гормонов (глюкагона, катехоламинов, тираксина, трийодтиранина) приводит к активации ферментов фосфорилазы и глюкозо-6-фосфатазы в гепатоцитах и соответственно к усилению мобилизации гликогена из депо.
Таким образом, при нарушених гормонального баланса, в частности при различных формах сахарного диабета (первичного и вторичного), возникает несостоятельность глюкостатической функции печени. При этом глюкоза не утилизируется в инсулинзависимых органах и тканях, а также в печени, несмотря на то, что проницаемость мембран гепатоцитов при вышеуказанных гормональных нарушениях для глюкозы не меняется. Однако в условиях дефицита инсулина блокируются все последующие метаболические пути ее превращения. Последнее приводит к развитию гипергликемии, глюкозурии, лактат- и/или кетоацидоза жировой инфильтрации печени и к тяжелым морфофункциональными изменениям со стороны различных органов и систем.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом