
Развитие костной ткани на месте хряща протекает несколько сложнее, чем остеогистогенез, совершающийся непосредственно в мезенхиме. В этом случае развитию костной ткани предшествует образование хрящевой модели трубчатой кости, выполняющей опорную функцию на докостной стадии формирования скелета. Исходными клетками являются камбиальные клетки надхрящницы — адвентициальные. При подрастании к надхрящнице кровеносных сосудов и улучшении условий трофики и оксигенации эти клетки дифференцируются не в хондробласты, а в остеобласты, вырабатывающие межклеточное вещество ретикулофиброзной костной ткани. Они образуют подобие костной манжетки, окружающей хрящевую модель будущей трубчатой кости. Так возникает перихондральная костная ткань и надкостница. Окруженные костной тканью хрящевые клетки, утратившие связь с источником питания, подвергаются дегенерации. В возникшие полости дегенерирующего хряща из надкостницы врастают кровеносные сосуды с расположенными вокруг них камбиальными клетками. Некоторые из них превращаются в остеобласты, обусловливающие энхондральное развитие ретикулофиброзной костной ткани. Клетки, которые замуровываются в межклеточное вещество, дифференцируются в остеоциты, а периферически расположенные клетки — остеобласты — размножаются и продолжают синтез и секрецию компонентов межклеточного вещества. Все эти процессы первоначально протекают в середине хрящевой модели трубчатой кости (диафизе) и распространяются в проксимальном и дистальном направлениях.
В зоне контакта хрящевой и костной тканей можно выделить зоны неизмененного хряща, размножающихся хондроцитов, формирующих клеточные колонки, зону дегенерации и замещения хряща костной тканью. Зона размножающихся хрящевых клеток определяет зону роста будущей кости и важна для формирования вектора роста кости.
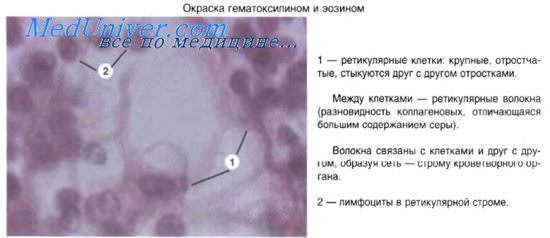
Одновременно с формированием ретикулофиброзной костной ткани, содержащей остеобласты и остеоциты, возникает другой гистогенетический тип клеток — остеокласты. Это крупные многоядерные (до 20-100 ядер) клетки размером до 100 мкм в диаметре являются производными стволовой кроветворной клетки. Цитоплазма остеокластов оксифильна со слабо развитой эндоплазматической сетью. Хорошо развит комплекс Гольджи. В цитоплазме много лизосом, содержащих кислую фосфатазу, коллагеназу, карбоангидразу и другие ферменты. Особенно много лизосом в той части цитоплазмы остеокластов, которая обращена к разрушаемой ткани. На этой поверхности имеются многочисленные выросты цитоплазмы, образующие подобие «щеточной (гофрированной) каемки». Остеокласты специализированы на «внеклеточной работе» лизосом: гидролитические ферменты из них выходят и резорбируют межклеточное вещество. Методами микрокиносъемки показано, что остеокласты подвергают деминерализации и разрушению оссеиновые волокна и аморфное вещество, а затем макрофаги фагоцитируют остатки органического субстрата. Остеокласты разрушают хрящевую ткань и ретикулофиброзную костную ткань, формируя каналы для врастающих сосудов и проникновения остеобластов.
Последующие стадии гистогенеза складываются из процессов новообразования костной ткани, ее разрушения остеокластами и перестройки — ремоделирования. Важным фактором гистогенеза пластинчатой костной ткани, входящей в состав трубчатой кости, является вектор роста кости. Он определяет направление движения остеокластов, следовательно, формирования каналов и врастание в них кровеносных сосудов (по вектору роста). Кровеносный сосуд, в свою очередь, определяет упорядоченное (концентрическое) расположение остеобластов вокруг себя. При этом остеобласты синтезируют межклеточное вещество, оссеиновые волокна которого упорядоченно (параллельно) располагаются возле остеобласта и при минерализации формируют костную пластинку, толщиной 3-10 мкм. Соседняя костная пластинка содержит оссеиновые фибриллы, которые располагаются под углом по отношению к первым.
На протяжении гистогенеза и всей возрастной динамики костной ткани в ней происходит непрерывная перестройка благодаря согласованной деятельности остеобластов и остеоцитов, образующих межклеточное вещество, а также остеокластов, разрушающих костную ткань, что необходимо для процессов ее самообновления. Так происходит смена генераций костных пластинок и формирующихся структурно-функциональных единиц — остеонов, достигается упорядоченность расположения последних, следовательно, высокая механическая прочность костной ткани и кости как органа (см. кость).
Дентиноидная костная ткань отличается отсутствием тел костных клеток в толще межклеточного вещества. Дентин — это вещество, состоящее из коллагеновых волокон и основного аморфного вещества, пропитанного минеральными солями. Образующие дентин зуба клетки — одонтобласты (точнее — их ядросодержащая часть) — расположены вне дентина в пульпе зуба. Дентин пронизан дентинными канальцами, в которых проходят отростки одонтобластов. Сходное строение имеет цемент зуба.
Ретикулофиброзная (грубоволокнистая) костная ткань характеризуется беспорядочным расположением оссеиновых фибрилл в виде толстых, плотных пучков волокон и основного аморфного вещества. Такая костная ткань образует кости в зародышевом и раннем постнатальном периодах. У взрослого человека она сохраняется лишь на месте прикрепления сухожилии к кости, в зарастающих швах черепа, а также в составе тканевого регенерата на месте переломов костей.
Пластинчатая костная ткань отличается упорядоченным расположением оссеиновых фибрилл в составе костных пластинок. Последние образуют расположенные один за другим слои пропитанного солями кальция фибрилл, образованных остеобластами. Слои имеют толщину от 3-7 до нескольких сотен микрометров. Каждая костная пластинка состоит из параллельно ориентированных тонких оссеиновых (коллагеновых) волокон (коллаген 1-го типа). Но коллагеновые волокна двух прилежащих друг к другу костных пластинок ориентированы под разными углами. Костная пластинка соединяется с соседней пластинкой коллагеновыми фибриллами. Так создается прочная волокнистая основа кости. Костные пластинки располагаются концентрически вокруг сосудов, то есть формируют остеоны — структурно-функциональные единицы пластинчатой кости как органа. Кроме этого существуют наружные и внутренние окружающие и вставочные пластинки трубчатой кости (см. ниже).
Регенерация. В регенерации костной ткани участвуют детерминированные остеогенные элементы в составе надкостницы, механоциты костного мозга, которые размножаются и дифференцируются в остеобласты. Продуцируя межклеточное вещество, остеобласты дифференцируются в остеоциты и образуют ретикулофиброзную костную ткань. Кроме того, адвентициальные клетки волокнистой соединительной ткани надкостницы также принимают участие в регенерации костной ткани. Однако дифференцировка их во многом зависит от микроокружения, внетканевых и внеорганных факторов (например, от репозиции отломков, неподвижности отломков, оксигенации места перелома и др.).
Дифференцировка адвентициальных клеток возможна в трех направлениях: остеогенном, хондрогенном, фибробластическом. Этим определяется соотношение различных видов тканей в регенерате. При преимущественно остеобластическом гистогенезе формируется ретикулофиброзная костная ткань, которая постепенно ремоделируется с образованием костной ткани, напоминающей по своему строению пластинчатую.
Вопрос 13: Костные ткани. Классификация. Диффероны костной ткани (см и эм). Характеристика матрикса. Механизм минерализации и резорбции.
Костные ткани образуют скелет, защищающий внутренние органы от повреждений, входящий в локомоторный аппарат и являющийся важнейшим депо минеральных веществ в организме.
Классификация костных тканей основана на различиях строения межклеточного вещества, в частности, степени упорядоченности расположения в нем коллагеновых волокон. Выделяют (1) грубоволокнистую костную ткань и (2) пластинчатую костную ткань. Грубоволокнистая (ретикулофиброзная) костная ткань характеризуется неупорядоченным расположением коллагеновых волокон в матриксе. Она отличается относительно небольшой механической прочностью и обычно образуется тогда, когда остеобласты формируют остеоид с высокой скоростью. Лакуны с телами остеоцитов не имеют закономерной ориентации. Содержание остеоцитов в грубоволокнистой костной ткани выше, чем в пластинчатой, а в ее матриксе больше основного вещества и меньше минеральных компонентов. В ходе нормального развития и при регенерации костной ткани грубоволокнистая костная ткань постепенно замещается пластинчатой. Пластинчатая костная ткань у взрослого образует практически весь костный скелет. Ее минерализованное межклеточное вещество состоит из особых костных пластинок толщиной 3-10 мкм, каждая из которых содержит параллельно расположенные тонкие коллагеновые волокна. Волокна соседних пластинок лежат под углом друг к другу, что способствует равномерному распределению действующих на них нагрузок. Пластинки в кости образуют нескольких систем. Лакуны, содержащие тела остеоцитов, располагаются между пластинками упорядоченно, а костные канальцы, в которых находятся отростки клеток, пронизывают пластинки под прямыми углами. Минерализация органического матрикса остеобластами осуществляется двумя основными механизмами: (1) путем отложения кристаллов гидроксиапатита из перенасыщенной внеклеточной жидкости вдоль фибрилл коллагена. Секретируемые остеобластами неколлагеновые белки контролируют ход минерализации. В частности, костный сиалопротеин и остеонектин усиливают связывание минеральных веществ и регулируют рост кристаллов гидроксиапатита. Особую роль в процессах формирования начального ядра отложения кристаллов приписывают некоторым протеогликанам, занимающим зоны зазоров между молекулами тропоколлагена в коллагеновых фибриллах. Эти протеогликаны связывают кальций, удерживая его в зонах зазоров; в дальнейшем они разрушаются ферментами, а с коллагеном в области зазоров связываются фосфопротеины. Их фосфат реагирует с ионами кальция, образуя первые кристаллы минералов. Процесс протекает с участием щелочной фосфатазы, обеспечивающей дефосфорилирование и локальное повышение концентраций фосфатных ионов. (2) посредством секреции особых матричных пузырьков — мелких (100-200 нм) округлых мембранных структур, которые образуются и выделяются в матрикс остеобластами. Эти пузырьки содержат высокие концентрации фосфата кальция и щелочной фосфатазы, им свойственна высокая активность других мембранных ферментов и липидов. Микросреда внутри матричных пузырьков способствует отложению первых кристаллов гидроксиапатита. В этом процессе важную роль приписывают щелочная фосфатазе, отщепляющей фосфат, связанный с органическими веществами, который далее участвует в образовании кристаллов гидроксиапатита. Разрушаясь, пузырьки служат ядрами, вокруг которых растут кристаллы гидроксиапатита. В дальнейшем очаги минерализации увеличиваются в размерах и сливаются друг с другом, превращая новообразованный остеоид в зрелый костный матрикс.
Читайте также: Каким клеем клеить джинсовую ткань
Механизм резорбции костной ткани остеокластами. 1) прикрепление остеокластов к резорбируемой поверхности кости обеспечивается рядом адгезивных взаимодействий, опосредованных интегринами и белками матрикса. При этом в остеокласте наблюдается выраженная перестройка элементов цитоскелета. 2) закисление содержимого лакун осуществляется двумя механизмами: (а) путем выделения кислого содержимого вакуолей в лакуну; (б) благодаря действию протонных насосов, накачивающих ионы Н+ в лакуну; 3) резорбцию минерального компонента матрикса, которая осуществляется вследствие воздействия на него кислого содержимого лакуны; 4) растворение органических компонентов матрикса вследствие действия лизосомальных ферментов остеокластов, секретированных ими в лакуну и активирующихся при низких значениях pH. Высказывается мнение о том, что остеокласты осуществляют лишь деминерализацию матрикса, а разрушение органических компонентов обеспечивается макрофагами; 5) удаление продуктов разрушения костной ткани осуществляется двумя механизмами: (а) их утечкой из лакуны после отделения плазмолеммы от поверхности кости, (б) поглощением продуктов остеокластами, и их везикулярным транспортом через цитоплазму клетки с последующим выделением в области ее апикального полюса.
Дифферон остеоцита: стволовые и полустволовые клетки, остеогенные клетки, остеобласты, остеоциты. Клетки образуются из малодифференцированных клеток мезенхимы; у взрослых стволовые и полустволовые клетки имеются во внутреннем слое надкостницы, во время образования кости находятся на ее поверхности и вокруг внутрикостных сосудов. Остеокласт. Дифферон остеокласта включает дифферон моноцита, затем моноцит выходит из кровеносного русла и трансформируется в макрофаг. Несколько макрофагов сливаются, и образуется многоядерный симпласт –остеокласт. В остеокласте много ядер, большой объём цитоплазмы.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Остеогенный дифферон костной ткани

Особенности метаболизма, клинико-молекулярные механизмы ремоделирования костной ткани и регуляция этих процессов в последние годы привлекает пристальное внимание. Это связано с тем, что остеопороз в начале ХХІ столетия стал одним из наиболее распространенных заболеваний, наряду с онкологическими процессами, сахарным диабетом и сердечно-сосудистой патологией, занимает ведущее место в структуре заболеваемости и смертности населения [17]. В Германии (82 млн. жителей) остеопорозом (ОП) страдает до 7,8 млн. жителей страны старше 50-ти лет [16]. В России количество больных ОП составляет около 14 млн. человек [22]. Частота выявления ОП в Европе у женщин достигает 36 %, у мужчин – 26,4 % [14]. Клиническими последствиями остеопороза являются переломы позвонков, трубчатых костей, ребер. В России каждые 5 минут происходит перелом шейки бедра, вызванный ОП, а в течение года в стране происходит 9 млн. переломов периферического скелета и более 3 млн. переломов позвонков [5]. Перелом проксимального отдела бедра является причиной смерти 14-49 % пациентов в течение первого года после травмы, и этот показатель значительно выше у мужчин, чем у женщин [2,14]. Половина больных, выживших после перелома бедра, нуждается в постоянном длительном уходе из-за снижения качества жизни. Важной медицинской проблемой является остеопенический синдром, развивающийся вследствие других заболеваний: эндокринных, ревматологических, онкологических, болезней органов пищеварения, почек, легких, как осложнения при приеме некоторых медикаментозных средств: иммунодепрессантов, глюкокортикостероидов, тиреоидных гормонов и др. [7]. Значительное распространение ОП и остеопоротических переломов среди населения, тяжесть исходов, существенные затраты на лечение и реабилитацию больных отражают высокую социальную значимость заболевания.
Кость – специализированная соединительная ткань, содержащая минерализованную внеклеточную фазу, которая позволяет выполнять опорные и метаболические функции. Основными клетками костной ткани являются остеоциты, остеобласты и остеокласты. Клетки костной ткани характеризуются высокой метаболической активностью и имеют четкое разделение функций. Особенностью метаболизма костной ткани является ее перестройка на протяжении всей жизни, поскольку в отличие от других тканей кость обновляется не только заменой «старых» макромолекул вновь синтезируемыми, но реформируется и на морфологическом уровне. Перестройка костной ткани характеризуется двумя понятиями: моделированием и ремоделированием.
Моделирование определяет характерную форму микроструктуры кости в процессе ее роста, восстанавливает кость при переломах, перестраивая костную мозоль и адаптируя ее при заживлении. Активация процессов моделирования осуществляется под влиянием метаболических и механических факторов и сводится к пространственной координации процессов резорбции и формирования кости, происходящих одновременно в различных участках ткани.
Процесс ремоделирования заключается в полном разрушении точечных участков кости (резорбции) и заполнении возникающих дефектов новообразованной костью (костеобразование). Оба эти процесса тесно взаимосвязаны и являются результатом клеточного взаимодействия остеокластов (ОК) и остеобластов (ОБ). В детском и юношеском возрасте превалирует остеогенез и костная масса возрастает на 8 % в год. После 40 лет процесс резорбции начинает преобладать над костеобразованием, в результате масса и прочность кости постепенно снижаются. У взрослого человека результаты ремоделирования сбалансированы и это позволяет сохранять постоянство массы кости. В кортикальной кости костный обмен протекает в более медленном темпе, в трабекулярной кости – более интенсивно. В трубчатых костях ремоделирование осуществляется на трех поверхностях: периостальной, эндоостальной, к которой относится и поверхность губчатого вещества и в системе гаверсовых каналов, а в теле позвонка – только на периостальной и эндостальной. На поверхности периоста в течение всей жизни сохраняется положительный баланс перестройки, на поверхности гаверсовых каналов перестройка уравновешена, а на эндостальной поверхности доминирует отрицательный баланс. Это обуславливает истончение кортикального слоя и рарефекацию губчатой кости.
В процессе ремоделирования участвуют ОК, ОБ, остеоциты, активные мезенхимальные клетки. ОБ – берут начало от мезенхимальных стволовых клеток, ОК – от макрофагально-моноцитарных клеток костного мозга. ОБ – мононуклеарная клетка, обеспечивающая процесс остеогенеза, характеризуется развитыми субклеточными структурами, отвечающими за биосинтетические процессы, и обилием митохондрий. По мере образования компонентов остеоида и минерализации вокруг себя ОБ снижают биосинтетические процессы и трансформируются в остеоциты. ОК – гигантская многоядерная клетка, осуществляющая резорбцию, т.е. рассасывание костной ткани, действуя только на минерализованную кость. ОК отличаются высокой концентрацией лизосом, содержащих набор кислых гидролаз, участвующих в расщеплении макромолекул остеоида, характеризуются высокой активностью Н+-АТФ-азы, карбоангидразы, а также способностью выделять в среду изофермент кислой фосфатазы, не ингибирующейся при действии тартрата. Остеоциты, ОБ, преостеобласты продуцируют молекулы внеклеточного матрикса; адгезивные молекулы на поверхности клеток, обеспечивающие контакты межклеточные и с молекулами внеклеточного матрикса; ростовые факторы и их антагонисты, регулирующие обновление и дифференцировку клеток.
Читайте также: Как красиво обернуть букет тканью
Внеклеточный матрикс кости по белковому составу близок к собственно соединительной ткани. Его фибриллярные структуры примерно на 90 % состоят из коллагена І типа, содержит минорные коллагены V и XII типов. Коллагены придают прочность, эластичность костной ткани, поддерживают адгезию, пролиферацию и дифференциацию клеток с остеобластным фенотипом, участвуют в процессах минерализации и др. Коллаген V типа связывается с протеогликанами (особенно гепарансульфатными), а также молекулами тромбоспондина и других белков межклеточного матрикса. Коллаген XII типа также ассоциируется с протеогликанами (особенно хондротинсульфатными), имеет участки подобия к фибронектину, содержит несколько центров связывания с клетками (последовательность Арг-Гли—Асп), участвует в функционально-структурном единении межклеточных структур с клеточными элементами кости, опосредуя прикрепление клеток к волокнам коллагенов типа І и V, и взаимодействует с неколлагеновыми протеинами [6].
Костный матрикс содержит большое разнообразие неколлагеновых белков, представленные гликопротеинами, фосфопротеинами и протеогликанами. Среди них остеокальцин, фибронектин, остеопонтин, ламинин, виментин, костный сиалопротеин II, матриксный Gla-протеин, остеопонтин, декорин, диглан и др. Одни из них являются адгезивными белками (фибронектин, ламинин, остеонектин), другие выполняют специфические функции: остеокальцин – кальций связывающий и кальций транспортирующий белок, прочно связанный с гидроксиопатитом и участвующий в реализации кальциевых эффектов Д-гормона; костный сиалопротеин II, остеопонтин – основные нуклеаторы в процессе минерализации внеклеточного матрикса; остеонектин и матриксный Gla-протеин-регуляторы минерализации костного матрикса [6].
В цикле ремоделирования различают фазы активации (инициации), резорбции, реверсии, формирования (остеогенеза) и покоя. Фундаментальную роль в инициации костного ремоделирования и регуляции метаболической активности клеток костной ткани играют ОБ. Инициация ремоделирования осуществляется в местах нарушения микроструктуры кости, постоянно происходящих в процессе жизнедеятельности. Остеопонтин и остеокальцин эндотелиальной мембраны и матрикса кости стимулируют рекрутирование предшественников ОК к локусу дефекта кости и дифференцировку до зрелых ОК. Таким образом, активация и регуляция ремоделирования костной ткани является следствием взаимодействия между ОБ и ОК [23]. Ключевую роль в формировании, дифференцировке и активности ОК играет цитокиновая система RANKL-RANK-OPG [8]. RANKL-гликопротеин, который продуцируется клетками остеобластного ряда и активированными Т-лимфоцитами, является основным стимулом для созревания ОК. RANKL, экспрессированный на поверхности ОБ, связывается с RANK. RANK – рецептор, расположенный на плазматической мембране предшественников ОК. Он, приводя к внутриклеточным каскадным механизмам, воздействует на ядерный фактор каппа-B (NF-kB). NF-kB с помощью рецептора TRAF 6 поступает из цитоплазмы в ядро и повышает экспрессию протеина NFATс1, являющийся специфическим триггером, запускающим процесс транскрипции внутриклеточных генов, формирующих процесс остекластогенеза [13,24]. ОБ и стволовые мезенхимальные клетки костного мозга одновременно синтезируют макрофагально-колонийстимулирующий фактор (M-CSF), который, связываясь со своим высоко аффинным трансмембранным рецептором (c-fms), активирует внутриклеточную тирозинкиназу, также стимулирующую процесс пролиферации и дифференциации клеток-предшественниц ОК [19]. OPG – остеопротегерин – растворимый рецептор для RANKL, синтезируется клетками остеобластного фенотипа, а также β-лимфоцитами, клетками стромы и эндотелия сосудов. OPG является блокатором взаимодействия RANKL с RANK и, как следствие, угнетает формирование ОК и резорбцию костной ткани [19, 24]. При действии на остеобласты паратиреоидного гормона, кальцитриола, интерлейкинов 1 и 6 (ИЛ-1, ИЛ-6), фактора некроза опухоли (TNF) пролиферативная активность M-CSF значительно возрастает, под влиянием эстрагенов и OPG понижается [24]. Глюкокортикостероиды увеличивают в ОБ экспрессию RANKL, изменяют соотношение RANKL и OPG, и это приводит к увеличению остеокластогенеза [25].
Дифференцированный ОК занимает определенное положение на поверхности клетки и конструирует специализированный цитоскелет, позволяющий ему создать изолированную полость резорбции – микросреду между костью и ОК. При активации ОК экспрессируются avb интегрины – адгезивные трансмембранные рецепторы клеточной поверхности, вступающие во взаимодействие с коллагеном І типа, остеопонтином, сиалопротеином и другими белками внеклеточного матрикса, содержащими центр связывания с клетками (Арг-Гли-Асп последовательности). При этом интегриновый рецептор индуцирует в цитоплазме ОК повышение уровня Са2+ и рН, а также фосфорилирование ряда протеинов по тирозину, которые контролируют контакт ОК с внеклеточным матриксом. Особую роль среди них играет тирозиновая протеинкиназа, сопряженная с цитоплазматическим доменом β-субъединицы интегрина. Последующее фосфорилирование по тирозину ряда цитоплазматических белков ОК включает цепь последовательной передачи сигналов другим молекулам: G-протеинам, цитоплазматическим протеинкиназам и транскрипционным факторам клеточного ядра с экспрессией генов ОК, ответственным за продукцию компонентов резорбирующей активности клетки [8].
В фазе резорбции плазматическая мембрана ОК, обращенная к изолированной кости, формирует множество складок, многократно увеличивая резорбирующую поверхность, образует гофрированную резорбтивную мембрану. В микросреду созданной полости резорбции ОК выделяет протоны Н+ с помощью плазматической Н+ -АТФазы. Образование Н+ катализируется карбоангидразой ІІ, а внутриклеточная рН поддерживается путем обмена ионами НСО3-/Cl-. Анионы HCO3- выводятся в межклеточную среду, а Cl- поступают в клетку и по анионным каналам гофрированной мембраны выводятся в микрополость. рН в резорбтивной полости снижается до 4-4,5. Создаются условия для растворения кристаллов гидроксиапатита минеральной фазы кости и деградации органического матрикса кислыми гидролитическими ферментами, включая катепсин К, которые с помощью микровезикул высвобождаются в полость резорбции. Синтез и накопление катепсина К модулируется факторами, оказывающими влияние на функцию ОК, – TNF, ИЛ-1, RANKL, эстрогенами, простагландином Е2 [15]. Фосфатные группы ряда неколлагеновх белков в зоне резорбции отщепляет кислая фосфатаза (тартрат-резистентная). Определенную роль в деградации пептидных цепей играет и супероксидный анион-радикал, активно генерируемые в зоне резорбции. Продукты резорбции минеральной фазы и остеоида удаляются путем трансцитоза мембранных везикул остеокластов и механизмом «разгерметизации» изолированной полости. В результате действия ОК в кости образуется лакуна резорбции [6] – в кортикальной кости появляются конусовидные пустоты, в губчатой кости – углубления, имеющие форму блюдца. Продолжительность фазы резорбции 10-12 суток. Она завершается переходным периодом – фазой реверсии.
Фаза реверсии представляет финал резорбции. В ней происходит апоптоз ОК, привлечение остеогенных клеток, их пролиферация и дифференцировка в зрелые ОБ. Регуляция этими процессами в переходной фазе осуществляется факторами локальной (межклеточной и внутриклеточной) регуляции, образующихся и функционирующих в пределах костной ткани. Это группа ростовых факторов внеклеточного матрикса и молекулы внутриклеточной сигнальных систем (транскрипционные факторы, активирующие гены, ответственные за остеогенную дифференциацию клеток). Трансформирующий фактор роста β (TGF-β) способствует апоптозу ОК и хемотаксису преостеобластов и ОБ. Хемотаксису преостеобластов способствуют и остеокальцин, и фрагменты коллагена І типа. Остеокальцин также способствует реализации кальциевых эффектов кальцитриола, что характеризует его роль в дифференциации клеток-предшественниц остеобластов [6]. Дифференциация остеогенных клеток и регуляция функции ОБ осуществляется и другими факторами роста (фактор роста фибриногенобластов, инсулиноподобный фактор роста, β-катенин, костные морфогенетические белки и др.), гормонами (паратгормон, кальцитриол и др.) вызывающими экспрессию генов транскрипционных факторов, контролирующих остеосинтез: Сbfa -1 (core binding factor alpha-1), известный как RUNX-2 (runt related transcription factor-2); Twist; Osterix/Sp 7; Dlx 5; Msx-2; NF-kB, Варх 1 (смотри подробнее 3, 24). Важнейшим из них является Сbfa -1, который непосредственно регулирует функции многих генов, участвующих в образовании белков костной ткани: коллагена типа І, остеокальцина, остеопонтина, матриксной металлопротеазы 1, костного сиалопротеина, щелочной фосфатазы, RANKL, С/ЕВР, рецептора TGF- β [20]. Для активации RUNX-2 остеогенных белков необходима его кооперация с костным морфогенетическим белком-2 (ВМР-2), который стимулирует ацетилирование RUNX-2 гистонацетилтрансферазой, повышая ее стабильность и активность, подавляет его деацетилирование гистондеацетилазами и деградацию RUNX-2, опосредуемую убиквитинлигазой Smurf-1. В результате активность RUNX-2 контролируется динамикой равновесия процессов его ацетилирования, деацетилирования и убиквитинизации [12].
Читайте также: Комплект для ткани ворота
Для дифференциации преостеобластов в зрелые ОБ необходимо участие транскрипционного фактора Osterix/Sp7, так же как и RUNX-2. Osterix/Sp7 вызывает экспрессию генов коллагена типа І, костного сиалопротеина, остеопонтина, остеонектина, остеокальцина. Osterix/Sp7 экспрессируется под влиянием гена Dlx 5, активируемого ВМР-2. Dlx 5 может также активировать и экспрессию RUNX-2. Dlx 5 является ключевым белком созревания ОБ [12].
Остеогенную дифференциацию снижают транскрипционные факторы Twist, способные угнетать связывание с ДНК и активацию гена RUNX-2 в предшественниках ОБ [18]. NF-kB регулируют большую группу генов, участвующих в клеточном росте и клеточной адгезии. При остеогенезе транскрипционный фактор NF-kB снижает дифференциацию ОБ, контролируя kB участки промотора гена RUNX-2 [3].
Две группы воздействия влияют на дифференцировку мезенхимальных стволовых клеток ОБ – химические сигналы и физическое напряжение цитоскелета клеток, которое активирует в них сигнальные пути [3]. Важнейшими регуляторами остеогенной дифференциации являются секреторные белки семейства Wnt: суперсемейство трансформирующее рост фактора β1,- β2,- β3, активины, ингибины; факторы роста фибробластов -2,-9; инсулиноподобный фактор роста [1,3,20], костные морфогенетические белки. ВМР-2,4,7 – стимулирует дифференциацию клеток остеобластного ряда в остеоциты. Они связываются с рецепторами на клетках-предшественницах ОБ, с участием трансмембранного белка неогенина индуцируют фосфорилирование специализированных белков цитоплазмы Smad -1, -5,-8, которые передают сигналы в ядро клетки, активирующие экспрессию остеогенных белков. ВМР могут активировать и каскад киназ, стимулирующий р38 митогенактивирующие протеинкиназы. Оба пути сигнализации работают синергично, индуцируют экспрессию транскрипционных факторов Dlx 5, RUNX-2 и Oсterix в дифференцирующихся ОБ, сопровождающееся повышенной продукцией белков костного матрикса [3].
В формировании костного скелета, в регуляции остеобластогенеза и функции ОБ важную роль играет канонический винглес-бета-катенин (Wnt/β-catenin) сигнальный путь [1,21]. Во время остеогенной дифференцировки в клетках резко увеличивается ряд лигандов Wnt (Wnt-2,4,5,11,16) с тропным к ним Frizzled-рецепторным комплексом (трансмембранный белок Frizzled) и сопряженные с ним ко-рецепторы липопротеиннов низкой плотности (LRP 5 и 6). Активация Wnt рецепторного комплекса приводит к усилению функции белка Disheveled, который ингибирует связанные с ним протеины GSK-3, APC и AXIN. Снижение активности киназы гликогенсинтетазы-3 (GSK-3) стабилизирует β-катенин, способствует его накоплению в цитоплазме и транслокации в ядро клетки, где β-катенин вступает во взаимодействие с транскрипционными факторами TCF/LEF/RUNX 2 (TCF – фактор внутриядерной транскрипции генов, LEF – лимфоидный фактор, повышающий процесс связывания внутриядерных компонентов) и регулирует экспрессию генов, ответственных за стимуляцию регенерации костной ткани, включая синтез циклина Д1, обеспечивающего продвижение клетки по клеточному циклу. Без активации Wnt корецепторов LRP 5 и LRP 6 β-катенин быстро разрушается. Wnt-лиганды взаимодействуют с TCF-β, потенцируют остеогенные эффекты ВМР-2. Экспрессию TGF-β и ВМР-2 в остеобластах, усиливают действие фактора роста фибробластов-2,-9, вызывая синергичный эффект [3].
При интенсивном образовании кости наблюдается увеличение в плазме крови паратиреоидного (ПТГ), соматропного (СТГ) гормонов и кальцитриола. СТГ активирует синтез инсулиноподобного фактора роста (IGP-1), протеогликанов и коллагенов в костной ткани, кальцийтриол – ее минерализацию [11]. Синтез IGP-1 в ОБ контролирует также ПТГ, который уменьшает старение и апоптоз ОБ, индуцирует угнетение экспрессии белков-ингибиторов циклин – зависимых киназ – Р21 и Р16, поддерживает дифференциацию остеогенных клеток [10].
Механическая активация остеогенных клеток связана с напряжением физической связи β1-интегринов преостеобластов, ОБ с белками внеклеточного матрикса (коллаген, ламинин, фибронектин и др.), восприятием напряжения внутриклеточными актином и миозином, с включением сигнальной передачи к ядру клетки, вызывая экспрессию транскрипционного фактора с –Fos и рост продукции IGP-1, интерлейкина – 8, простагландинов, стимулирующих остеогенез [3].
Регуляция остеогенеза имеет и механизмы подавления остеобластов. Его реализация осуществляется через Wnt/β – катенин сигнальный путь. Внеклеточные антагонисты этого пути ингибируют остеобластогенез связыванием Wnt белков активаторов (LRP 5 и LRP 6, Wnt – ингибирующий фактор 1) или образованием комплекса с одним из компонентов Wnt-рецептора – Dikkopf-1 и склеростин. При этом происходит быстрое разрушение убиквитин-протеосомальным механизмом β-катенина в цитоплазме, наблюдается снижение Wnt – сигнализации и уменьшение роста кости. Активация экспрессии генов склеростина и Dikkopf-1 в ОБ происходит при интенсивной активации RUNX 2 и других генов ВМР, способной привести к образованию избыточной костной массы, являясь, таким образом, контуром отрицательной обратной связи [1]. Дифференциация ОБ тормозится и ВМР-3, а также молекулами-антагонистами ВМР -2,-4,-5,-6,-7, секретируемыми ОБ в интерцеллюлярный матрикс кости – ногтином, хордином, филистатином и др. [12]. Паратиреоидный гормон, с одной стороны, тормозит экспрессию склеростина, а с другой, имеется и механизм предупреждения образования костной массы под воздействием ПТГ – активированные рецепторы 2-го типа TGF-β снижают активность рецептора -1 ПТГ [12].
Стадия формирования кости связана с биосинтетической функцией ОБ, секретирующие во внеклеточное пространство коллагены, неколлагеновые белки, ферменты и формирующие остеоид, который через 10-15 суток при активном участии ОБ начинает минерализовываться [6]. Каждый ОБ синтезирует и наращивает вокруг себя новый костный матрикс, минерализует его и превращается в остеоцит с отростками в системе канальцев, связывающим его с соседними клетками. Совокупность остеоцитов, отростки которых по костным каналам проникают в окружающие их костные пластинки, образует единую сеть в кости. Каждый остеоцит контактирует с соседними клетками и неактивными ОБ. Благодаря этому, поверхность обмена кости очень велика – 10 см3 кости имеет поверхность до 330 м2 [6].
Длительность цикла ремоделирования колеблется от 6 до 9 месяцев. Скорость обмена скелета в год составляет около 10 %, при этом скорость обмена кортикальной кости (85 % скелета) – 4 %, трабекулярной кости (15 % скелета) – 28 % в год [4]. Ремоделирование, во-первых, позволяет изменить объем, форму и плотность кости, максимально соответствуя существующим нагрузкам, поддерживая, корректируя и обновляя микроархитектонику ткани; во-вторых, является частью системы, обеспечивающей кругооборот некоторых важнейших минеральных соединений – Ca, Mg, P и др. в организме и сохранения их оптимальной концентрации в биологических средах.
Заключая обзор, можно резюмировать, что формирование, рост, развитие, ремоделирование, функционирование и метаболизм костной ткани осуществляется сложным взаимодействием костных клеток и нескольких групп регуляторов, включающих локальные факторы, в том числе продуцирующие самими костными клетками: TGF- β, простагландины, интерлейкины, ВМР и др.; системные ростовые факторы: макрофагальный колониестимулирующий фактор, гранулоцитарно-макрофагальный колониестимулирующий фактор, инсулиноподобные факторы роста, фибробластный фактор роста и др.; кальций регулирующие гормоны – ПТГ, кальцитонин, кальцитриол; другие системные гормоны: СТГ, инсулин, глюкокортикоиды, йодированные тиронины, глюкокортикоиды, половые. Факторы роста, цитокины, молекулы адгезии, синтезируемые в костном мозге и костных клетках, обеспечивают взаимодействие между клетками и матриксом кости, между самими клетками, опосредуют эффекты механических сдвигов, эффекты системных гормонов.
Бутолин Е.Г., д.м.н., профессор, заведующий кафедрой биологической химии ГБОУ ВПО «Ижевская государственная медицинская академия» Министерства здравоохранения Российской Федерации, г. Ижевск;
Лунева С.Н., д.б.н., профессор, руководитель научно-клинического диагностического отдела ФГБУ «Российский научный центр «Восстановительная травматология и ортопедия» им. ак. Г.А. Илизарова» МЗ РФ, г. Курган.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом