
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

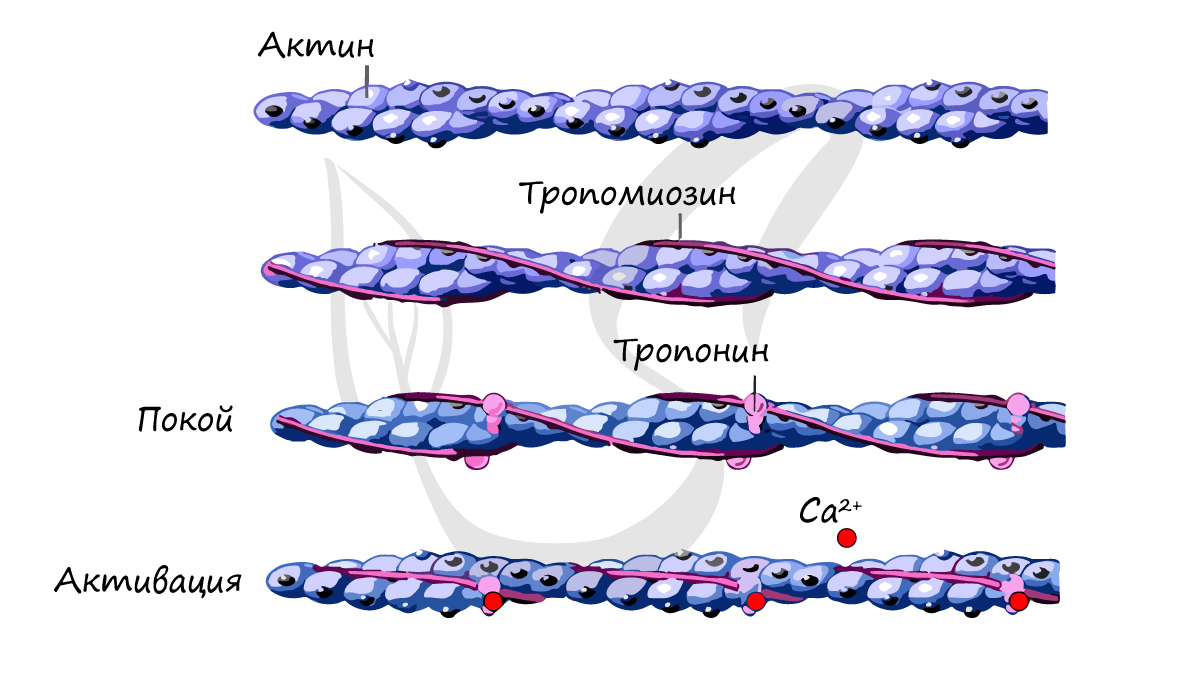
Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Читайте также: Секреторная ткань человека это
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Поперечно полосатая сердечная мышечная ткань скорость сокращения
Тема: Физиология мышечной ткани
I . Классификация мышц. Строение и свойства поперечно-полосатых мышц.
Перемещение тела в пространстве, поддержание определенной позы, работа сердца и сосудов и пищеварительного тракта у человека и позвоночных животных осуществляются мышцами двух основных типов: поперечнополосатыми (скелетной, сердечной) и гладкими, которые отличаются друг от друга клеточной и тканевой организацией, иннервацией и в определенной степени механизмами функционирования. В то же время в молекулярных механизмах мышечного сокращения между этими типами мышц есть много общего.
Различают 2 типа мышечных волокон:
1. Поперечнополосатые (скелетные и сердечные)
На рис. 12 схематически изображено поперечнополосатое мышечное волокно с основными структурами, обеспечивающими его возбуждение и сокращение. К этим структурам относятся:
1) Поверхностная мембрана (сарколемма), образующая продольные углубления – Т-трубочки.
2) Саркоплазматический ретикулум, служащий депо Са 2+ .
3) Миофибрилла – пучки параллельных нитей актина и миозина.
Скелетная мышца представляет собой сложную систему, преобразующую химическую энергию в механическую работу и тепло. В настоящее время хорошо исследованы молекулярные механизмы этого преобразования.
Структурная организация мышечного волокна . Мышечное волокно является многоядерной структурой, окруженной мембраной и содержащей специализированный сократительный аппарат — миофибриллы. Кроме этого, важнейшими компонентами мышечного волокна являются митохондрии, системы продольных трубочек — саркоплазматическая сеть (ретикулум) и система поперечных трубочек — Т-система. Функциональной единицей сократительного аппарата мышечной клетки является саркомер; из саркомеров состоит миофибрилла. Саркомеры отделяются друг от друга Z-пластинками. Саркомеры в миофибрилле расположены последовательно, поэтому сокращение саркомеров вызывает сокращение миофибриллы и общее укорочение мышечного волокна.


Рис. 11. Виды мышечной ткани

Рис. 12. Структура скелетных мышц
А — организация цилиндрических волокон в скелетной мышце, прикрепленной к костям сухожилиями. Б — структурная организация филаментов в волокне скелетной мышцы, создающая картину поперечных полос. Показаны многочисленные миофибриллы в одиночном мышечном волокне, а также организация толстых и тонких филаментов в саркомере
Поперечнополосатое мышечное волокно состоит из актиновых и миозиновых нитей. Миозиновые нити обладают двойным лучепреломлением (анизотропией), поэтому они образуют толстые нити (А-диски, анизатропные), актиновые нити обладают одинарным лучепреломлением (изотропией) и образуют тонкие нити ( I -диски, изотропные). Актиновые нити располагаются в промежутках между миозиновыми.
При электронной микроскопии видно, что на боковых сторонах миозиновой нити обнаруживаются выступы, получившие название поперечных мостиков. Они ориентированы по отношению к оси миозиновой нити под углом 120 0 . Согласно современным представлениям, поперечный мостик состоит из головки и шейки. Головка приобретает выраженную АТФ-азную активность при связывании с актином. Шейка обладает эластическими свойствами и представляет собой шарнирное соединение, поэтому головка поперечного мостика может поворачиваться вокруг своей оси.
На головке имеются участок связывания с актином и участок связывания с АТФ.
Актиновая нить состоит из трех белков: 1) двух скрученных цепочек актина, 2) тпропомиозина, который пронизывает цепочки актина и 3) тропонина, который «сидит» на цепочках актина и блокирует актиновую нить.
Посередине актиновых нитей проходит Z –линия. Участок, ограниченный двумя Z –линиями образует саркомер (функциональная единица сократительного аппарата). Таким образом, саркомер состоит из 2х половинок дисков I и одного целого А-диска.
Возбуждение, возникающее в одном мышечном волокне скелетной мышцы, распространяется только в пределах данного волокна и не переходит на соседние волокна. Следовательно, целая скелетная мышца подчиняется закону силы: при слабых раздражениях возбуждается и сокращается меньшее количество волокон, а при сильных – большее. Это увеличение количества сокращающихся волокон при увеличении силы раздражителя, называется пространственной суммацией, или вовлечением.
Существует два типа мышечных волокон:
Белые мышцы обеспечивают быстрые короткие сокращения, красные – более медленные длительные сокращения. Цвет красных волокон обусловлен большим содержанием миоглобина, который связывает и запасает кислород, необходимый для длительного сокращения. Также в красных мышцах больше содержания митохондрий и более густая сеть капилляров.
Любая мышца содержит оба типа волокон, но в какой-то мышце тот или иной тип преобладает. Например, в быстрых глазодвигательных мышцах содержатся в основном белые волокна, а в медленных мышцах спины – красные.
Читайте также: Как сделать кошелек из ткани своими руками 2 класс
Преобладание белых или красных мышц генетически предопределено. Эту особенность используют современные тренера при отборе детей в те или иные секции спорта. Если у ребенка преобладают белые волокна, то его лучше брать в секции, где требуется быстрота движений (например, бег на короткие дистанции), если же преобладают красные волокна – в секции, где требуется выносливость (например, марафонские бега).
Типичная гладкомышечная клетка изображена на рис. 13. Основные структурные черты, обусловливающие ее физиологические особенности следующие:
1) Вместо упорядоченных миофибрилл со строгим чередованием нитей актина и миозина имеются пучки актиновых нитей, в центре которых располагается миозин.
2) Эти пучки не располагаются параллельно оси гладкомышечной клетки, но прикрепляются к ее мембране в области так называемых плотных телец и при сокращении как бы сморщивают клетку.
3) Саркоплазматический ретикулум развит слабо.
В большинстве органов гладкие мышцы состоят из тесно связанных между собой клеток, образующих мышечные пласты. Клетки в пределах этих пластов соединены межклеточными контактами – щелевыми контактами (нексусами), позволяющими импульсу перескакивать от одной клетки к другой. Благодаря этому пласты гладкомышечных клеток возбуждаются и сокращаются как единое целое, подчиняясь закону «все или ничего».
Гладкие мышцы иннервируются волокнами вегетативной нервной системы, а не соматической, как скелетные мышцы.
Нервные волокна не образуют на гладкомышечных клетках типичных синапсов: нервные окончания разветвляются над мышечным пластом, причем между нервными волокнами и мышечными клетками остается довольно большое пространство. В ветвлениях нервных окончаний имеются варикозные расширения, в которых скапливаются пузырьки с медиатором; при возбуждении нервного окончания медиатор высвобождается из всех варикозных расширений, изливаясь на большую поверхность гладкомышечного пласта.
Исключение составляют мышцы зрачка, ресничного тела и некоторые другие гладкие мышцы, от которых требуется относительно быстрые и точные сокращения. Они состоят из отдельных мышечных волокон, разделенных прослойками межклеточного вещества и сокращающихся независимо друг от друга.
Гладкие мышцы, как и скелетные, обладают возбудимостью, проводимостью и сократимостью. Однако они отличаются по этим свойствам от поперечнополосатых.
У гладкомышечных клеток ПП менее отрицателен, чем у нейронов и скелетных мышц: обычно он составляет 0т -50 до -60 мВ. Основная причина этого — относительно более высокая проницаемость для натрия в покое.
Быстрых натриевых каналов, отвечающих за ПД в нейронах и скелетных мышцах, в гладкомышечных клетках практически нет. Следовательно ПД в этих клетках формируется за счет другого входящего тока, а именно – входа Са 2+ по кальциевым каналам. Эти каналы в отличие от натриевых, пропускают преимущественно Са 2+ и их ворота срабатывают значительно медленнее, т.е. медленнее открываются и медленнее закрываются.
Некоторые гладкомышечные клетки обладают свойством автоматизма, то есть способностью самопроизвольно генерировать ПД. У этих клеток нет потенциала покоя, но имеются, так называемы медленные волны, представляющие собой медленную самопроизвольную деполяризацию. Когда эта деполяризация достигает Екр, возникает один или несколько ПД.
Общие принципы сокращения едины и для гладких, и для скелетных мышц. В то же время сокращение гладких мышц отличается от сокращения скелетных. В скелетной мышце единственным источником кальция служит саркоплазматический ретикулум. В гладкой мышце он выражен слабо, поэтому главный источник – вход Са 2+ из внеклеточной среды по кальциевым каналам сарколеммы. Таким образом, вход Са 2+ по этим каналам играет двоякую роль: обеспечивает развитие ПД и запуск сокращения.
В гладкой мышце, как и в скелетной Са 2+ запускает взаимодействие актина с миозином через регуляторные белки. Однако, и сами эти белки и их действие совершенно иные.
1) Са 2+ связывается с белком кальмодулином.
2) Комплекс Са 2+ -кальмодулин активирует фермент киназу легких цепей миозина.
3) Этот фермент фосфорилирует миозин.
4) Только после этого поперечные мостики миозина приобретают способность связываться с активным центром актина.
Главное отличие в энергетике сокращения гладких мышц заключается в том, что миозин обладает слабым сродством к АТФ. Поэтому миозиновые мостики в течение длительного времени остаются связанными с актином. Это делает невозможными быстрые сокращения, но дает огромную экономию энергии, так как на образование каждого мостика тратится 1 молекула АТФ. В связи с этим, гладкие мышцы, уступая скелетным в скорости сокращения и расслабления, существенно выигрывают в экономичности и поэтому идеальны для поддержания длительных тонических сокращений внутренних органов.
Поперечнополосатую и гладкую мышцу можно сравнить с судном, которому приходиться удерживаться посередине реки против течения. Поперечнополосатая мышца подобна гребной лодке, которой надо постоянно работать веслами; это требует больших затрат энергии, но зато дает большую скорость и маневренность. Гладкая мышца – это плот, на котором от дна периодически отталкиваются шестом, затрат энергии при этом меньше, но быстрые движения невозможны.
Сердечная мышца занимает промежуточное положение между поперечнополосатыми и гладкими: по строению она ближе к п-п мышцам, по свойствам – к гладким. Сердечная мышца имеет поперечнополосатую исчерченность, однако, между волокнами имеются щелевые контакты с низкой электросопротивлением (нексусы). В связи с этим сердечная мышца, также как и гладкая, не обладает изолированным проведением возбуждения и подчиняется закону «все или ничего». Сердечная мышца обладает автоматией за счет наличия клеток, способных к самовозбуждению (пейсмекеров).
II . Молекулярные механизмы мышечного сокращения. Теория скольжения. Роль АТФ в механизмах мышечного сокращения.
Механизм мышечного сокращения . В процессе сокращения мышечного волокна в нем происходят следующие преобразования:
1.На сарколемме возникает ПД.
2. ПД проводится по сарколемме, что приводит к деполяризации Т-трубочек.
1. Деполяризация Т-трубочек приводит к открыванию кальциевых каналов саркоплазматического ретикулума.
2. Из саркоплазматического ретикулума выходит Са 2+ .
3. Са 2+ запускает взаимодействие актина с миозином, происходит их скольжение друг относительно друга и мышца сокращается.
4. Са 2+ закачивается обратно в саркоплазматический ретикулум с помощью Са 2+ — АТФ-азы (кальциевого насоса).
5. Взаимодействие актина с миозином прекращается, мышца расслабляется.
Са 2+ действует на актин и миозин не непосредственно, а через регуляторные белки. В поперечнополосатой мышце этими белками являются тропонин и тропомиозин.
Читайте также: Текстильные ткани в розницу
Регуляторные белки, сократительные белки и Са 2+ взаимодействуют следующим образом:
— в отсутствии Са 2+ активные центры нитей актина прикрыты нитями тропомиозина. С нитями тропомиозина связан тропонин (рис. 14);
— при поступлении к миофибриллам Са 2+ этот ион связывается с тропонином;
— взаимодействие Са 2+ с тропонином приводит к смещению нитей тропомиозина, активные центры открываются и становятся доступными для присоединения миозиновых поперечных мостиков.

Рис. 14. Молекулярная организация толстых и тонких филаментов: А — тонкий филамент, Б — молекула миозина, В — взаимодействие тонкого и толстого филамента
Из этого механизма следуют две важные особенности сокращения одиночного волокна скелетной мышцы:
1) Сила сокращения пропорциональна концентрации Са 2+ в цитоплазме (саркоплазме): чем больше эта концентрация, тем больше открывается активных центров на актине и больше к ним присоединяется миозиновых мостиков.
2) Сила сокращения одиночного мышечного волокна не зависит от силы раздражителя, то есть подчиняется закону «все или ничего». Это связано с тем, что одиночный раздражитель независимо от силы вызывает один ПД, а количество Са 2+ , выбрасываемого из саркоплазматического ретикулума в ответ на один ПД, всегда одинаково.
На один цикл поперечного мостика (связывание миозинового мостика с актином – гребковое движение – отсоединение) требуется одна молекула АТФ. Важно, что она присоединяется к мостику, когда он связан с актином, и только после этого мостик приобретает способность отсоединиться. В отсутствие АТФ миозиновые мостики постоянно связаны с актином.
Описанные механизмы укорочения мышечного волокна позволяют предположить, что для расслабления в первую очередь необходимо понижение концентрации ионов Са 2+ . Экспериментально было доказано, что саркоплазматическая сеть имеет специальный механизм – кальциевый насос, который активно возвращает кальций в цистерны.
Активация кальциевого насоса осуществляется неорганическим фосфатом, который образуется при гидролизе АТФ, а энергообеспечение работы кальциевого насоса также за счет энергии, образующейся при гидролизе АТФ. Таким образом, АТФ является вторым важнейшим фактором, абсолютно необходимым для процесса расслабления. Некоторое время после смерти мышцы остаются мягкими вследствие прекращения тонического влияния мотонейронов. Затем концентрация АТФ снижается ниже критического уровня и возможность разъединения головки миозина с актиновым филаментом исчезает. Возникает явление трупного окоченения с выраженной ригидностью скелетных мышц.
Феномен «защелки». Этот феномен заключается в том, что после развития напряжения гладкая мышца способна долго оставаться сокращенной, причем энергозатраты на это сокращение резко падают (мышца как бы «защелкивается» в сокращенном состоянии). Это важнейшая особенность гладких мышц, многие из которых часами и даже сутками пребывают в постоянном тонусе. Механизм этого эффекта до конца не понятен, ясно лишь, что в защелкнутом состоянии нити миозина теряют способность отсоединяться от актина.

Рис. 15. Механизм сокращения скелетной мышцы


Рис. 16. Сокращение гладкомышечной клетки
Пластичность. При повышении действующей на гладкую мышцу растягивающей силы эта мышца через какое-то время удлиняется, и ее сопряжение остается прежним. Например, при наполнении мочевого пузыря давление в нем сначала повышается, затем он растягивается, и давление в нем снова снижается. При постоянном растяжении гладкая мышца пребывает в «защелкнутом» состоянии, если растягивающая сила увеличивается, мышца удлиняется и мостики вновь «защелкиваются» в новом положении.
Утомление мышцы. Утомление – это временное снижение работоспособности. Существует несколько теорий мышечного утомления.
1. Накопление продуктов обмена (в частности, молочной и фосфатной кислот) (теория Э.Пфлюгера);
2. Теория истощения депо кальция, истощения энергетических запасов (гликоген, АТФ) (теория Шиффа).
В целом, в организме утомляются вначале нервные центры, затем синапсы и только после этого мышечные волокна.
III . Одиночное и тетаническое мышечное сокращение.
На кривой сокращения мышцы можно выделить три фазы (рис. 17):
1) Латентный период (от нанесения раздражителя до начала сокращения). Он включает время, необходимое для высвобождения Са 2+ , его диффузии к нитям актина, связывания с тропонином и пр.);

Рис. 17. Кривая одиночного мышечного сокращения
Повторный раздражитель, нанесенный на мышцу во время сокращения, способен вызвать новое возбуждение и сокращение. Это повторное сокращение складывается с предыдущим, то есть возникает временная суммация сокращений (сложение сокращений во времени). Суммация нескольких сокращений называется тетанусом.
Если повторный раздражитель попадает в фазу расслабления предшествующего сокращения, то возникает зубчатый тетанус (рис. 18), если в фазу укорочения – гладкий тетанус. Таким образом, гладкий тетанус возникает в ответ на действие раздражителя большей частоты, чем зубчатый.

Рис. 18. Виды сокращений мышечной ткани
Суммация и тетанус обусловлены тем, что при частых повторных раздражениях Са 2+ не успевает закачиваться обратно в саркоплазматический ретикулум и накапливается в цитоплазме, не позволяя мышце расслабиться и вызывая все более сильное сокращение.
Мышечный тонус. Мышцы человека не бывают полностью расслаблены, они всегда в состоянии некоторого напряжения, называемого мышечным тонусом. При этом медленные двигательные единицы сокращаются с небольшой частотой и поддерживают определенное положение тела в пространстве — позу, необходимую для осуществления фазических кратковременных движений. Мышечный тонус доставляет большие затруднения хирургам. После перелома бедра необходимо обеспечить вытяжение ноги, чтобы кости срослись конец в конец. Без вытяжения под действием мышечного тонуса кости срастутся неправильно, что приведет к укорочению ноги.
Сила мышц. Мышца при своем сокращении способна поднять большой груз, масса которого во много раз превышает массу самой мышцы.
Сила мышцы измеряется тем максимальным грузом, который она в состоянии поднять. Зависит сила мышцы от числа мышечных волокон, составляющих данную мышцу, и толщины этих волокон; она прямо пропорциональная физиологическому поперечному сечению, т. е. сумме попeречных сечений всех входящих в нее волокон. В мышцах с продольно расположенными волокнами физиологическое поперечное сечение совпадает с анатомическим — площадью поперечного разреза мышцы, проведенного перпендикулярно ее длине. В перистых и косых мышцах физиологическое сечение больше и соответственно больше мышечная сила.
При физической тренировке происходит утолщение мышечных волокон и увеличиваются их энергетические ресурсы. В связи с этим возрастает сила мышц.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом