
Поперечно полосатые мышечные ткани механизм сокращения
Выделяют несколько последовательных этапов запуска и осуществления мышечного сокращения.
1. Потенциал действия распространяется вдоль двигательного нервного волокна до его окончаний на мышечных волокнах.
2. Каждое нервное окончание секретирует небольшое количество нейромедиатора ацетилхолина.
3. Ацетилхолин действует на ограниченную область мембраны мышечного волокна, открывая многочисленные управляемые ацетилхолином каналы, проходящие сквозь белковые молекулы, встроенные в мембрану.
4. Открытие управляемых ацетилхолином каналов позволяет большому количеству ионов натрия диффундировать внутрь мышечного волокна, что ведет к возникновению на мембране потенциала действия.
5. Потенциал действия проводится вдоль мембраны мышечного волокна так же, как и по мембране нервного волокна.
6. Потенциал действия деполяризует мышечную мембрану, и большая часть возникающего при этом электричества течет через центр мышечного волокна. Это ведет к выделению из саркоплазматического ретикулума большого количества ионов кальция, которые в нем хранятся.
7. Ионы кальция инициируют силы сцепления между актиновыми и миозиновыми нитями, вызывающие скольжение их относительно друг друга, что и составляет основу процесса сокращения мыщц.
8. Спустя долю секунды с помощью кальциевого насоса в мембране саркоплазматического ретикулума ионы кальция закачиваются обратно и сохраняются в ретикулуме до прихода нового потенциала действия. Удаление ионов кальция от миофибрилл ведет к прекращению мышечного сокращения.
Далее мы обсудим молекулярные механизмы этого процесса.
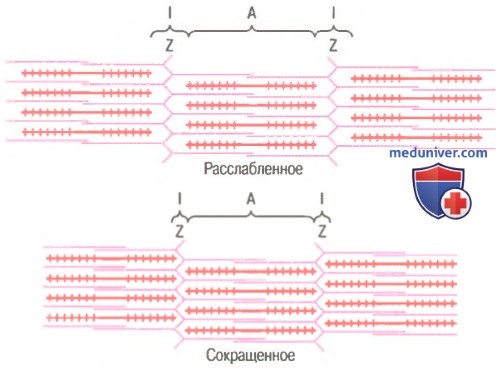
Миофибрилла в расслабленном и сокращенном состоянии. Показано (вверху), что актиновые нити (розовые) вдвинуты в пространства между миозиновыми нитями (красные). Сближение Z-дисков друг с другом (внизу).
Молекулярные механизмы мышечного сокращения
Механизм скольжения нитей для мышечного сокращения. На рисунке показан основной механизм мышечного сокращения. Показано расслабленное состояние саркомера (вверху) и сокращенное состояние (внизу). В расслабленном состоянии концы актиновых нитей, отходящие от двух последовательных Z-дисков, лишь незначительно перекрываются. Наоборот, в сокращенном состоянии актиновые нити втягиваются внутрь между миозиновыми так сильно, что их концы максимально перекрывают друг друга. При этом Z-диски притягиваются актиновыми нитями к концам миозиновых. Таким образом, мышечное сокращение осуществляется путем механизма скольжения нитей.
Что заставляет нити актина скользить внутрь среди нитей миозина? Это связано с действием сил, генерируемых при взаимодействии поперечных мостиков, исходящих от нитей миозина, с нитями актина. В условиях покоя эти силы не проявляются, однако распространение потенциала действия вдоль мышечного волокна приводит к выделению из саркоплазматическо-го ретикулума большого количества ионов кальция, которые быстро окружают миофи-бриллы. В свою очередь, ионы кальция активируют силы взаимодействия между нитями актина и миозина, в результате начинается сокращение. Для осуществления процесса сокращения необходима энергия. Ее источником являются высокоэнергетические связи молекулы АТФ, которая разрушается до АДФ с высвобождением энергии. В следующих разделах мы приведем известные детали молекулярных процессов сокращения.
Молекулярные особенности сократительных нитей
Миозиновая нить. Она состоит из множества молекул миозина, молекулярная масса каждой составляет около 480000. На рисунке показана отдельная молекула; и также — объединение многих молекул миозина в миозиновую нить, а также взаимодействие одной стороны этой нити с концами двух актиновых нитей.
В состав молекулы миозина входят 6 полипептидных цепей: 2 тяжелые цепи с молекулярной массой около 200000 каждая и 4 легкие цепи с молекулярной массой около 20000 каждая. Две тяжелые цепи спирально закручиваются вокруг друг друга, формируя двойную спираль, которую называют миозиновым хвостом. С одного конца обе цепи изгибаются в противоположных направлениях, формируя глобулярную полипептидную структуру, называемую миозиновой головкой. Таким образом, на одном конце двойной спирали молекулы миозина образуются 2 свободные головки; 4 легкие цепи также включены в состав миозиновой головки (по 2 в каждой). Они помогают регулировать функцию головки во время мышечного сокращения.
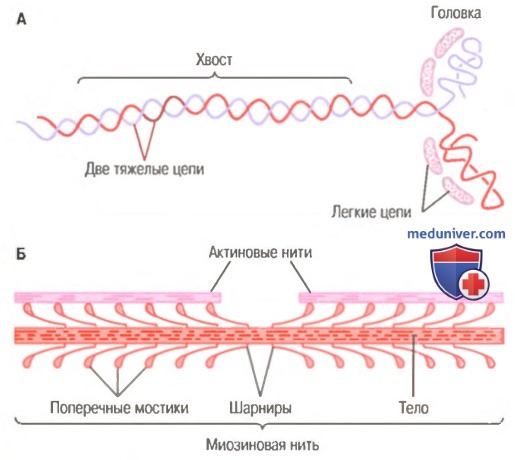
А. Молекула миозина.
Б. Объединение многих молекул миозина в одну миозиновую нить.
Показаны также тысячи миозиновых поперечных мостиков и взаимодействие их головок с прилежащими актиновыми нитями.
Миозиновая нить состоит из 200 или более отдельных молекул миозина. Видно, что хвосты молекул миозина объединяются, формируя тело нити, а многочисленные головки молекул выдаются наружу по сторонам тела. Кроме того, наряду с головкой в сторону выступает часть хвоста каждой миозиновой молекулы, образуя плечОу которое выдвигает головку наружу от тела, как показано на рисунке. Выступающие плечи и головки вместе называют поперечными мостиками. Каждый поперечный мостик может сгибаться в двух точках, называемых шарнирами. Один из них расположен в месте, где плечо отходит от тела миозиновой нити, а другой — где головка крепится к плечу. Движение плеча позволяет головке или выдвигаться далеко наружу от тела миозиновой нити, или приближаться к телу. В свою очередь, повороты головки участвуют в процессе сокращения, что обсуждается в следующих разделах.
Читайте также: Расчет ткани для постельного белья таблица выкройки
Общая длина каждой миозиновой нити остается постоянной и равна почти 1,6 мкм. В самом центре миозиновой нити на протяжении 0,2 мкм поперечных мостиков нет, поскольку снабженные шарнирами плечи отходят в стороны от центра.
Сама миозиновая нить сплетена таким образом, что каждая последующая пара поперечных мостиков смещена в продольном направлении относительно предыдущей на 120°, что обеспечивает распределение поперечных мостиков во всех направлениях вокруг нити.
АТФ-азная активность миозиновой головки. Есть и другая особенность миозиновой головки, необходимая для мышечного сокращения: миозиновая головка функционирует как фермент АТФ-аза. Как объясняется далее, это свойство позволяет головке расщеплять АТФ и использовать энергию расщепления высокоэнергетической связи для процесса сокращения.
Актиновая нить. Актиновая нить состоит из трех белковых компонентов: актина, тропомиозина и тропонина.
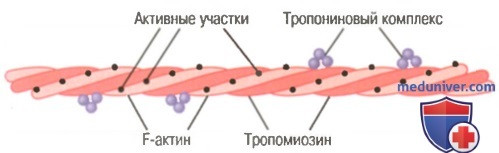
Актиновая нить, состоящая из двух спиралевидных цепочек молекул F-актина и двух цепочек молекул тропомиозина, расположенных в желобках между цепочками актина.
К одному концу каждой молекулы тропомиозина прикреплен тропониновый комплекс, который запускает сокращение.
Основой актиновой нити являются две цепи белковой молекулы F-актина. Обе цепи закручиваются в спираль так же, как и молекула миозина.
Каждая цепь двойной спирали F-актина состоит из полимеризованных молекул G-актина с молекулярной массой около 42000. К каждой молекуле G-актина прикреплена 1 молекула АДФ. Полагают, что эти молекулы АДФ являются активными участками на актиновых нитях, с которыми взаимодействуют поперечные мостики миозиновых нитей, обеспечивая мышечное сокращение. Активные участки на обеих цепях F-актина двойной спирали расположены со смещением таким образом, что вдоль всей поверхности актиновой нити встречается один активный участок примерно через каждые 2,7 нм.
Длина каждой актиновой нити — около 1 мкм. Основания актиновых нитей прочно встроены в Z-диски; концы этих нитей выступают в обоих направлениях, располагаясь в пространствах между миозиновыми молекулами.
Молекулы тропомиозина. Актиновая нить также содержит другой белок — тропомиозин. Каждая молекула тропомиозина имеет молекулярную массу 70000 и длину 40 нм. Эти молекулы спирально оплетают спираль из F-актина. В состоянии покоя молекулы тропомиозина располагаются поверх активных участков актиновых нитей, препятствуя их взаимодействию с миозиновыми нитями, лежащему в основе сокращения.
Тропонин и его роль в мышечном сокращении. По ходу молекул тропомиозина к ним периодически прикреплены другие белковые молекулы, называемые тропонином. Они представляют собой комплексы трех слабосвязанных белковых субъединиц, каждая из которых играет специфическую роль в регуляции мышечного сокращения. Одна из субъединиц (тропонин I) имеет высокое сродство к актину, другая (тропонин Т) — к тропомиозину, третья (тропонин С) — к ионам кальция. Считают, что этот комплекс прикрепляет тропомиозин к актину. Высокое сродство тропонина к ионам кальция, как полагают, инициирует процесс сокращения, о чем говорится в следующей статье.
Видео физиология мышц и мышечного сокращения — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Поперечно полосатые мышечные ткани механизм сокращения
Мышечное сокращение является жизненно важной функцией организма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений – ходьба, мимика, движения глазных яблок, глотание, дыхание и т. п. осуществляются за счет скелетных мышц. Непроизвольные движения (кроме сокращения сердца) – перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря – обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Структурная организация скелетной мышцы
Мышечное волокно и миофибрилла (рис. 1). Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл , которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.
Рис. 1. Иерархические уровни организации скелетной мышцы (по Bloom , 1968).
Саркомер и филламенты. Саркомеры в миофибрилле отделены друг от друга Z -пластинками, которые содержат белок бета-актинин. В обоих направлениях от Z -пластинки отходят тонкие актиновые филламенты. В промежутках между ними располагаются более толстые миозиновые филламенты .
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула белка актина . В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина , соединенные с нитевидными молекулами белка тропомиозина.
Миозиновые филламенты образованы повторяющимися молекулами белка миозина . Каждая молекула миозина имеет головку и хвост . Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик .
Клеточная мембрана мышечного волокна образует инвагинации (поперечные трубочки), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++ .
Двигательная единица. Функциональной единицей скелетной мышцы является двигательная единица (ДЕ). ДЕ – совокупность мышечных волокон, которые иннервируются отростками одного мотонейрона. Возбуждение и сокращение волокон, входящих в состав одной ДЕ, происходит одновременно (при возбуждении соответствующего мотонейрона). Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга.
Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий.
Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 ° . За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ (рис. 2, В).
Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается (рис. 2, Г).
Рис. 2. Механизм мышечного сокращения.
Объяснение – в тексте.
1 – актиновый филламент,
2 – центр связывания,
3 – миозиновый филламент,
4 – головка миозина,
5 – Z -диск саркомера.
Сопряжение возбуждения и сокращения в скелетной мышце
В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий.
Рис. 3. Механизм сопряжения возбуждения и сокращения.
Объяснение – в тексте.
1 – поперечная трубочка саркоплазматичекой мембраны,
2 –саркоплазматичекий ретикулум,
3 – ион Са++ ,
4 – молекула тропонина,
5 – молекула тропомиозина.
В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану.
Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В).
Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г).
К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д).
Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения .
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++ ; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А):
латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
фаза укорочения (около 50 мс);
фаза расслабления (около 50 мс).
Рис. 4. Характеристика одиночного мышечного сокращения. Происхождение зубчатого и гладкого тетануса.
Б – фазы и периоды иышечного сокращения,
Б – режимы мышечного сокращения, возникающие при разной частоте стимуляции мышцы.
Изменение длины мышцы показано синим цветом, потенциал действия в мышце — красным, возбудиумость мышцы — фиолетовым.
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 4, Б).
Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 4, A), в результате чего амплитуда тетануса значительно уменьшается.
Работа скелетной мышцы
Сила сокращения скелетной мышцы определяется 2 факторами:
числом ДЕ, участвующих в сокращении;
частотой сокращения мышечных волокон.
Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения.
Виды работы скелетной мышцы:
динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве;
статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении;
динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела.
Во время выполнения работы мышца может сокращаться:
изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы;
ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках.
Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмом мышечной работы утомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов.
Структурная организация и сокращение гладких мышц
Структурная организация . Гладкая мышца состоит из одиночных клеток веретенообразной формы (миоцитов), которые располагаются в мышце более или менее хаотично. Сократительные филламенты расположены нерегулярно, вследствие чего отсутствует поперечная исчерченность мышцы.
Механизм сокращения аналогичен таковому в скелетной мышце, но скорость скольжения филламентов и скорость гидролиза АТФ в 100–1000 раз ниже, чем в скелетной мускулатуре.
Механизм сопряжения возбуждения и сокращения . При возбуждении клетки Cа++ поступает в цитоплазму миоцита не только из саркоплазматичекого ретикулума, но и из межклеточного пространства. Ионы Cа++ при участии белка кальмодулина активируют фермент (киназу миозина), который переносит фосфатную группу с АТФ на миозин. Головки фосфорилированного миозина приобретают способность присоединяться к актиновым филламентам.
Сокращение и расслабление гладких мышц . Скорость удаления ионов Са++ из саркоплазмы значительно меньше, чем в скелетной мышце, вследствие чего расслабление происходит очень медленно. Гладкие мышцы совершают длительные тонические сокращения и медленные ритмические движения. Вследствие невысокой интенсивности гидролиза АТФ гладкие мышцы оптимально приспособлены для длительного сокращения, не приводящего к утомлению и большим энергозатратам.
Физиологические свойства мышц
Общими физиологическими свойствами скелетных и гладких мышц являются возбудимость и сократимость . Сравнительная характеристика скелетных и гладких мышц приведена в табл. 6.1. Физиологические свойства и особенности сердечной мускулатуры рассматриваются в разделе «Физиологические механизмы гомеостаза».
Таблица 7.1.Сравнительная характеристика скелетных и гладких мышц
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом