
а) Электрические явления во время торможения нейрона. Влияние тормозных синапсов на постсинаптическую мембрану. Тормозной постсинаптический потенциал. Тормозные синапсы открывают в основном хлорные каналы, что позволяет ионам хлора легко проходить через мембрану. Чтобы понять, как тормозные синапсы тормозят постсинаптический нейрон, нужно вспомнить, что мы знаем о потенциале Нернста для ионов Сl-. Мы рассчитали, что он равен примерно -70 мВ. Этот потенциал отрицательнее, чем мембранный потенциал покоя нейрона, равный -65 мВ. Следовательно, открытие хлорных каналов будет способствовать движению отрицательно заряженных ионов Сl- из внеклеточной жидкости внутрь. Это сдвигает мембранный потенциал в направлении более отрицательных значений по сравнению с покоем приблизительно до уровня -70 мВ.
Открытие калиевых каналов позволяет положительно заряженным ионам К+ двигаться наружу, что приводит к большей отрицательности внутри клетки, чем в покое. Таким образом, оба события (вход ионов Сl- в клетку и выход ионов К+ из нее) увеличивают степень внутриклеточной отрицательности. Этот процесс называют гиперполяризацией. Увеличение отрицательности мембранного потенциала по сравнению с его внутриклеточным уровнем в покое тормозит нейрон, поэтому выход значений отрицательности за пределы исходного мембранного потенциала покоя называют тормозным постсинаптическим потенциалом (ТПСП).
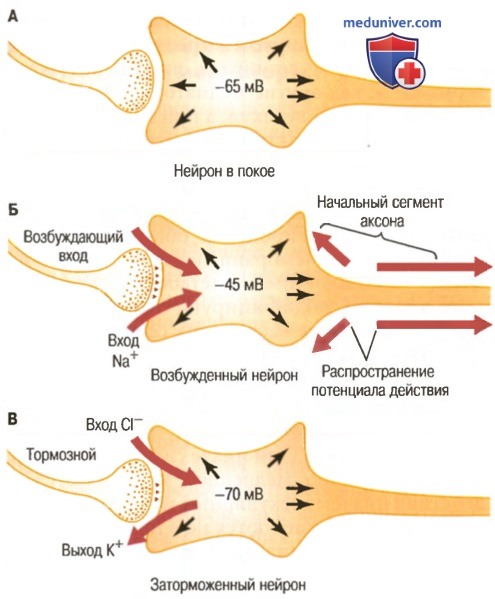
Три состояния нейрона.
А. Нейрон в покое с нормальным внутриклеточным потенциалом -65 мВ.
Б. Нейрон в состоянии возбуждения с менее отрицательным внутриклеточным потенциалом (-45 мВ), связанным с входом натрия.
В. Нейрон в заторможенном состоянии с более отрицательным внутриклеточным потенциалом (-70 мВ), что может быть связано с выходом ионов К+, входом ионов Cl- или с тем и другим
На рисунке В выше показано влияние активации тормозных синапсов, позволяющих ионам Сl-входить в клетку и/или ионам К+ выходить из нее, на мембранный потенциал. При этом он сдвигается от значения -65 мВ до более отрицательного значения -70 мВ. Этот мембранный потенциал на 5 мВ отрицательнее, чем в покое, и, следовательно, проведение нервного сигнала через синапс тормозит ТПСП, равный -5 мВ.
б) Пресинаптическое торможение. Кроме постсинаптического торможения, вызываемого тормозными синапсами, действующими на мембране нейрона, часто происходит торможение другого типа, которое развивается на пресинаптических терминалях, прежде чем сигнал достигает синапса. Этот тип торможения, называемый пресинаптическим торможением, осуществляется следующим путем.
Причиной пресинаптического торможения является выделение тормозного медиатора на наружную поверхность пресинаптических нервных волокон, прежде чем их собственные окончания достигнут поверхности постсинаптического нейрона. В большинстве случаев тормозным медиатором является ГАМК. При этом развивается специфический эффект, связанный с открытием анионных каналов, что позволяет большому числу ионов СГ диффундировать в терминальное волокно. Отрицательные заряды этих ионов тормозят синаптическое проведение, нейтрализуя большую часть возбуждающего эффекта положительно заряженных ионов Na+, которые также входят в терминальные волокна, когда здесь возникает потенциал действия.
Пресинаптическое торможение происходит во многих сенсорных путях нервной системы. Смежные сенсорные волокна часто взаимно тормозят друг друга, что сводит к минимуму боковое распространение и смешивание сигналов в чувствительных трактах.
Подробнее мы обсудили важность этого феномена в отдельных статьях на сайте — просим вас пользоваться формой поиска выше.
Видео физиология торможения в ЦНС — профессор, д.м.н. П.Е. Умрюхин
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Процесс торможения в тканях
Тип нервной системы имеет определенное значение в процессе приспособления к окружающей среде. Он определяет стрессоустойсивость, способность к обучению, время удержания внимания и многое другое, что было показано многочисленными работами, выполненными в школе И.П. Павлова [9, 10].
Я. Стреляу в своей работе «О темпераменте с точки зрения высшей нервной деятельности» разделяет позицию И.П. Павлова о том, что в основе темперамента лежат некоторые фундаментальные свойства нервных процессов – возбуждение и торможение. К этим свойствам относятся: сила возбуждения и торможения, их уравновешенность и подвижность [11].
И.П. Павлов различал силу процесса возбуждения и силу процесса торможения, считая их двумя независимыми свойствами нервной системы. Сила процесса возбуждения, с этой точки зрения, отражает работоспособность нервной клетки. Она проявляется в функциональной выносливости, способности выдерживать длительное или кратковременное, но сильное возбуждение. Мерой силы процесса возбуждения является способ реагирования на сильные, продолжительные или часто повторяющиеся раздражители [9].
Сила процесса торможения проявляется в способности к образованию различных тормозных условных реакций (например, угасание, дифференцировка или запаздывание). Оно связано с воздержанием от определенных действий или отсрочкой реакции. Чем адекватнее реакция в таких ситуациях, тем большей выносливостью относительно торможения обладает нервная система и, таким образом, тем сильнее процесс торможения [10]. Следовательно, торможение связано с волевыми процессами и, безусловно, представляется важнейшим параметром в процессе адаптации.
Подвижность нервных процессов, согласно И.П. Павлову, состоит в быстроте перехода одного нервного процесса в другой, «оба процесса должны, так сказать, поспевать за этими колебаниями, т. е. должны обладать высокой подвижностью, способностью быстро, по требованию внешних условий, уступать место, давать преимущество одному раздражению перед другим, раздражению перед торможением и обратно [9]. Подвижность нервных процессов обнаруживается в способности к изменению поведения при изменении внешней ситуации. Поэтому мерой этого свойства нервной системы является быстрота перехода от одного действия к другому, от пассивного состояния к активному, и наоборот [10].
Указанные свойства нервных процессов образуют определенные системы, комбинации; что и составляет тип нервной системы, или тип высшей нервной деятельности. Он складывается из характерной для отдельных индивидов совокупности основных свойств нервной системы – силы, уравновешенности и подвижности процессов возбуждения и торможения [10].
Соотношение свойств нервной системы может влиять на возможность адаптации человека к изменению внешних условий [2]. Есть данные о том, что люди с различной выраженностью латеральных признаков неодинаковым образом приспосабливаются к природным и социальным факторам среды [3, 4, 8, 14], и эти процессы протекают различным образом у мужчин и женщин [1]. Представляло интерес выявить возможную причину этих различий в особенностях свойств нервной системы людей с неодинаковой степенью выраженностью латеральных показателей.
Материалы и методы исследования
Было обследовано 175 молодых людей (25,4±2,7 года), из них 99 девушек (средний возраст 24,9±2,5 года) и 76 юношей (средний возраст 26,0±2,8 года). Все испытуемые на момент обследования обучались на курсах иностранного языка.
Читайте также: Возрастные изменения костной ткани кратко
Латеральные показатели и совокупный профиль функциональной сенсомоторной асимметрии (ФСМА) выявляли, описывая ведущую руку, ногу, ухо и глаз с помощью совокупности проб [6]. Пробы проводились трижды, в протокол вносились данные о каждом выполнении. Для выявления ведущей руки проводилось девять проб: теппинг-тест и восемь проб, предложенных В.П. Леутиным и Е.И. Николаевой [4]. «Переплетение пальцев рук», «Поза Наполеона», «Аплодирование», «Плечевой тест», «Рука, берущая предмет», «Рука, держащая ручку или карандаш», «Отвинчивание крышки на баночке», «Рисование круга и квадрата с закрытыми глазами».
Для определения ведущей ноги проводились девять проб [6]: «Положить ногу на ногу», «Подпрыгнуть на одной ноге», «Наступить на предмет», «Пнуть мяч ногой», «Движение с закрытыми глазами», «Шаг назад» (ведущей считается нога, первой начинающая движение), «Поставить колено на стул», «Прыжок вперед», «Наклон вперед с закрытыми глазами».
Ведущий глаз оценивался с помощью пяти проб: «Прицеливание» (осуществляется ведущим глазом), «Подмигивание» (ведущий глаз открыт), «Прищуривание», «Рассмотрение отдаленного предмета через полую трубу», «Дырка в карте» (действие осуществляет ведущий глаз).
Слуховая асимметрия оценивалась при помощи трех проб: «Тиканье часов», «Прислушивание к шуму на улице», «Шепотная речь».
Для оценивания типа профиля ФСМА высчитывались соответствующие общие показатели для ведущих руки, ноги, уха и глаза как среднее арифметическое по результатам выполнения проб. Затем вычислялось цифровое значение профиля путем суммирования соответствующих общих показателей и нахождения общего значения.
Обработка результатов проб проводилась при помощи метода, предложенного Е.Ю. Борисенковой и Е.И. Николаевой [7]. Каждому буквенному обозначению результата пробы присваивалось числовое значение: П – 2 балла, С – 1 балл, Л – 0 баллов, где П – выполнение пробы правой рукой, С – выполнение пробы двумя руками попеременно или одновременно, Л – выполнение пробы левой рукой [7]. Далее подсчитывался общий показатель ведущих руки, ноги, уха и глаза как среднее арифметическое по результатам выполнения проб. После этого проводилась проверка соответствия полученного распределения нормальному закону при помощи непараметрического критерия Колмогорова – Смирнова и программы обработки данных SPSS 17 [5].
Для определения темперамента и выраженности основных свойств нервной системы (процесса возбуждения, процесса торможения и подвижности нервных процессов) была использована методика изучения структуры темперамента Я. Стреляу [11] (адаптация Н.Н.Даниловой, А.Г.Шмелева).
Результаты исследования и их обсуждение
Анализ распределения испытуемых по типам профиля ФСМА обнаружил, что 10,3 % из них имели левый профиль, 55,4 % – смешанный и 34,4 % – правый. Эти данные соответствуют результатам других исследований [8,13], обнаруживших большее число людей со смешанными показателями профиля функциональной сенсомоторной асимметрии.
В табл.1 представлены данные теста Я Стреляу, направленного на описание темперамента, у юношей и девушек с различным типом профиля ФСМА. Нет различий в уровне возбуждения у испытуемых с разным типом профиля. Однако есть некоторая тенденция снижения этого уровня у девушек с правым профилем. Уровень возбуждения говорит о сильной ответной реакции на различные раздражители, быстрой включаемости в работу и более низкой утомляемости девушек с левым профилем.
Согласно полученным данным, и девушки, и юноши с левым профилем имеют самые низкие значения процессов торможения. Возможно, именно это объясняет высокую эмоциональную лабильность леворуких людей, отмеченную многими авторами 2.
У левопрофильных юношей по сравнению с левопрофильными девушками снижена подвижность нервных процессов. Таким образом, у девушек в целом более высокая переключаемость, мобильность нервных процессов и, следовательно, лучшие возможности для адаптации по сравнению с юношами. Самый низкий адаптационный потенциал отмечен у юношей с левым профилем по сравнению с юношами с другими латеральными признаками, хотя эти различия не значимы.
Показатели свойств нервной системы у юношей и девушек с различным типом профиля ФСМА
Торможение в ЦНС (И.М. Сеченов), его виды и роль. Современное представление о механизмах центрального торможения. Тормозные синапсы и их медиаторы. Ионные механизмы ТПСП
Торможение – активный процесс, возникающий при действии раздражителей на ткань, проявляется в подавлении другого возбуждения, функционального отправления ткани нет.
Торможение может развиваться только в форме локального ответа.
Выделяют два типа торможения:
1) первичное . Для его возникновения необходимо наличие специальных тормозных нейронов. Торможение возникает первично без предшествующего возбуждения под воздействием тормозного медиатора.
Различают два вида первичного торможения:
— пресинаптическое в аксо-аксональном синапсе;
— постсинаптическое в аксодендрическом синапсе.
2) вторичное . Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур, всегда связано с процессом возбуждения.
Виды вторичного торможения:
— запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона;
— пессимальное, возникающее при высокой частоте раздражения; парабиотическое, возникающее при сильно и длительно действующем раздражении;
— торможение вслед за возбуждением, возникающее вследствие снижения функционального состояния нейронов после возбуждения;
— торможение по принципу отрицательной индукции;
— торможение условных рефлексов.
Процессы возбуждения и торможения тесно связаны между собой, протекают одновременно и являются различными проявлениями единого процесса. Очаги возбуждения и торможения подвижны, охватывают большие или меньшие области нейронных популяций и могут быть более или менее выраженными. Возбуждение непременно сменяется торможением, и наоборот, т. е. между торможением и возбуждением существуют индукционные отношения.
Торможение лежит в основе координации движений, обеспечивает защиту центральных нейронов от перевозбуждения. Торможение в ЦНС может возникать при одновременном поступлении в спинной мозг нервных импульсов различной силы с нескольких раздражителей. Более сильное раздражение тормозит рефлексы, которые должны были наступать в ответ на более слабые.
В 1862 г. И. М. Сеченов открыл явление центрального торможения. Он доказал в своем опыте, что раздражение кристалликом хлорида натрия зрительных бугров лягушки (большие полушария головного мозга удалены) вызывает торможение рефлексов спинного мозга. После устранения раздражителя рефлекторная деятельность спинного мозга восстанавливалась. Результат этого опыта позволил И. М. Сеченому сделать заключение, что в ЦНС наряду с процессом возбуждения развивается процесс торможения, который способен угнетать рефлекторные акты организма. Н. Е. Введенский высказал предположение, что в основе явления торможения лежит принцип отрицательной индукции: более возбудимый участок в ЦНС тормозит активность менее возбудимых участков.
Читайте также: Ткань для футболок высокого качества
Современная трактовка опыта И. М. Сеченова (И. М. Сеченов раздражал ретикулярную формацию ствола мозга): возбуждение ретикулярной формации повышает активность тормозных нейронов спинного мозга – клеток Реншоу, что приводит к торможению α-мотонейронов спинного мозга и угнетает рефлекторную деятельность спинного мозга.
Тормозные синапсы образованы специальными тормозными нейронами (точнее, их аксонами). Медиатором могут быть глицин, ГАМК и ряд других веществ. Обычно глицин вырабатывается в синапсах, с помощью которых осуществляется постсинаптическое торможение. При взаимодействии глицина как медиатора с глициновыми рецепторами нейрона возникает гиперполяризация нейрона (ТПСП) и, как следствие, — снижение возбудимости нейрона вплоть до полной его рефрактерности. В результате этого возбуждающие воздействия, оказываемые через другие аксоны, становятся малоэффективными или неэффективными. Нейрон выключается из работы полностью.
Тормозные синапсы открывают в основном хлорные каналы, что позволяет ионам хлора легко проходить через мембрану. Чтобы понять, как тормозные синапсы тормозят постсинаптический нейрон, нужно вспомнить, что мы знаем о потенциале Нернста для ионов Сl-. Мы рассчитали, что он равен примерно -70 мВ. Этот потенциал отрицательнее, чем мембранный потенциал покоя нейрона, равный -65 мВ. Следовательно, открытие хлорных каналов будет способствовать движению отрицательно заряженных ионов Сl- из внеклеточной жидкости внутрь. Это сдвигает мембранный потенциал в направлении более отрицательных значений по сравнению с покоем приблизительно до уровня -70 мВ.
Открытие калиевых каналов позволяет положительно заряженным ионам К+ двигаться наружу, что приводит к большей отрицательности внутри клетки, чем в покое. Таким образом, оба события (вход ионов Сl- в клетку и выход ионов К+ из нее) увеличивают степень внутриклеточной отрицательности. Этот процесс называют гиперполяризацией. Увеличение отрицательности мембранного потенциала по сравнению с его внутриклеточным уровнем в покое тормозит нейрон, поэтому выход значений отрицательности за пределы исходного мембранного потенциала покоя называют ТПСП.
Функциональные особенности соматической и вегетативной нервной системы. Сравнительная характеристика симпатического, парасимпатического и метасимпатического отделов вегетативной нервной системы.
Первое и основное отличие строения ВНС от строения соматической состоит в расположении эфферентного (моторного) нейрона. В СНС вставочный и моторный нейроны располагаются в сером веществе СМ, в ВНС эффекторный нейрон вынесен на периферию, за пределы СМ, и лежит в одном из ганглиев — пара-, превертебральном или интраорганном. Более того, в метасимпатической части ВНС весь рефлекторный аппарат полностью находится в интрамуральных ганглиях и нервных сплетениях внутренних органов.
Второе отличие касается выхода нервных волокон из ЦНС. Соматические НВ покидают СМ сегментарно и перекрывают иннервацией не менее трех смежных сегментов. Волокна же ВНС выходят из трех участков ЦНС (ГМ, грудопоясничного и крестцового отделов СМ). Они иннервируют все органы и ткани без исключения. Большинство висцеральных систем имеет тройную (симпатическую, пара- и метасимпатическую) иннервацию.
Третье отличие касается иннервации органов соматической и ВНС. Перерезка у животных вентральных корешков СМ сопровождается полным перерождением всех соматических эфферентных волокон. Она не затрагивает дуги автономного рефлекса ввиду того, что ее эффекторный нейрон вынесен в пара- или превертебральный ганглий. В этих условиях эффекторный орган управляется импульсами данного нейрона. Именно это обстоятельство подчеркивает относительную автономию указанного отдела НС.
Четвертое отличие относится к свойствам нервных волокон. В ВНС они в большинстве своем безмякотные или тонкие мякотные, как, например, преганглионарные волокна, диаметр которых не превышает 5 мкм. Такие волокна принадлежат к типу В. Постганглионарные волокна еще тоньше, большая часть их лишена миелиновой оболочки, они относятся к типу С. В отличие от них соматические эфферентные волокна толстые, мякотные, диаметр их составляет 12-14 мкм. Кроме того, пре- и постганглионарные волокна отличаются низкой возбудимостью. Для вызова в них ответной реакции необходима значительно большая, чем для моторных соматических волокон, сила раздражения.
Волокна ВНС характеризуются большим рефрактерным периодом и большой хронаксией. Скорость распространения по ним НИ невелика и составляет в преганглионарных волокнах до 18 м/с, в постганглионарных — до 3 м/с. Потенциалы действия волокон ВНС характеризуются большей, чем в соматических эфферентах, длительностью. Их возникновение в преганглионарных волокнах сопровождается продолжительным следовым положительным потенциалом, в постганглионарных волокнах — следовым отрицательным потенциалом с последующей продолжительной следовой гиперполяризацией (300-400 мс).
ВНС обеспечивает экстраорганную и внутриорганную регуляцию функций организма и включает в себя три компонента:
Вегетативная нервная система обладает рядом анатомических и физиологических особенностей, которые определяют механизмы ее работы.
Анатомические свойства:
1. Трехкомпонентное очаговое расположение нервных центров. Низший уровень симпатического отдела представлен боковыми рогами с VII шейного по III–IV поясничные позвонки, а парасимпатического – крестцовыми сегментами и стволом мозга. Высшие подкорковые центры находятся на границе ядер гипоталамуса (симпатический отдел – задняя группа, а парасимпатический – передняя). Корковый уровень лежит в области шестого-восьмого полей Бродмана (мотосенсорная зона), в которых достигается точечная локализация поступающих нервных импульсов. За счет наличия такой структуры вегетативной нервной системы работа внутренних органов не доходит до порога нашего сознания.
2. Наличие вегетативных ганглиев. В симпатическом отделе они расположены либо по обеим сторонам вдоль позвоночника, либо входят в состав сплетений. Таким образом, дуга имеет короткий преганглионарный и длинный постганглионарный путь. Нейроны пара-симпатического отдела находятся вблизи рабочего органа или в его стенке, поэтому дуга имеет длинный преганглионарный и короткий постганглионарный путь.
3. Эффеторные волокна относятся к группе В и С.
Физиологические свойства:
1. Особенности функционирования вегетативных ганглиев. Наличие феномена мультипликации (одновременного протекания двух противоположных процессов – дивергенции и конвергенции). Дивергенция – расхождение нервных импульсов от тела одного нейрона на несколько постганглионарных волокон другого. Конвергенция – схождение на теле каждого постганглионарного нейрона импульсов от нескольких преганглионарных.
Это обеспечивает надежность передачи информации из ЦНС на рабочий орган. Увеличение продолжительности постсинаптического потенциала, наличие следовой гиперполяризации и синоптической задержки способствуют передаче возбуждения со скоростью 1,5–3,0 м/с. Однако импульсы частично гасятся или полностью блокируются в вегетативных ганглиях. Таким образом они регулируют поток информации из ЦНС. За счет этого свойства их называют вынесенными на периферию нервными центрами, а вегетативную нервную систему – автономной.
Читайте также: Халат из ткани капитоний
2. Особенности нервных волокон. Преганглионарные нервные волокна относятся к группе В и проводят возбуждение со скоростью 3—18 м/с, постганглионарные – к группе С. Они проводят возбуждение со скоростью 0,5–3,0 м/с. Так как эфферентный путь симпатического отдела представлен преганглионарными волокнами, а парасимпатического – постганглионарными, то скорость передачи импульсов выше у парасимпатической нервной системы.
Таким образом, вегетативная нервная система функционирует неодинаково, ее работа зависит от особенностей ганглиев и строения волокон.
Симпатическая нервная система осуществляет иннервацию всех органов и тканей (стимулирует работу сердца, увеличивает просвет дыхательных путей, тормозит секреторную, моторную и всасывательную активность желудочно-кишечного тракта и т. д.). Она выполняет гомеостатическую и адаптационно-трофическую функции.
Ее гомеостатическая роль заключается в поддержании постоянства внутренней среды организма в активном состоянии, т. е.симпатическая нервная система включается в работу только при физических нагрузках, эмоциональных реакциях, стрессах, болевых воздействий, кровопотерях.
Адаптационно-трофическая функция направлена на регуляцию интенсивности обменных процессов. Это обеспечивает приспособление организма к меняющимся условиям среды существования.
Таким образом, симпатический отдел начинает действовать в активном состоянии и обеспечивает работу органов и тканей.
Парасимпатическая нервная система является антагонистом симпатической и выполняет гомеостатическую и защитную функции, регулирует опорожнение полых органов.
Гомеостатическая роль носит восстановительный характер и действует в состоянии покоя. Это проявляется в виде уменьшения частоты и силы сердечных сокращений, стимуляции деятельности желудочно-кишечного тракта при уменьшении уровня глюкозы в крови и т. д.
Все защитные рефлексы избавляют организм от чужеродных частиц. Например, кашель очищает горло, чиханье освобождает носовые ходы, рвота приводит к удалению пищи и т. д.
Опорожнение полых органов происходит при повышении тонуса гладких мышц, входящих в состав стенки. Это приводит к поступлению нервных импульсов в ЦНС, где они обрабатывают и по эффекторному пути направляются до сфинктеров, вызывая их расслабление.
Метсимпатическая нервная система представляет собой совокупность микроганглиев, расположенных в ткани органов. Они состоят из трех видов нервных клеток – афферентных, эфферентных и вставочных, поэтому выполняют следующие функции:
— обеспечивает внутриорганную иннервацию;
— являются промежуточным звеном между тканью и экстраорганной нервной системой. При действии слабого раздражителя активируется метсимпатический отдел, и все решается на местном уровне. При поступлении сильных импульсов они передаются через парасимпатический и симпатический отделы к центральным ганглиям, где происходит их обработка.
Метсимпатическая нервная система регулирует работу гладких мышц, входящих в состав большинства органов желудочно-кишечного тракта, миокарда, секреторную активность, местные иммунологические реакции и др.
Роль СМ в процессах регуляции деятельности ОДА и вегетативных функций организма. Характеристика спинальных животных. Принципы работы спинного мозга. Клинически важные спинальные рефлексы.
СМ – наиболее древнее образование ЦНС. Характерная особенность строения – сегментарность.
Нейроны СМ образуют его серое вещество в виде передних и задних рогов. Они выполняют рефлекторную функцию СМ.
Задние рога содержат нейроны (интернейроны), которые передают импульсы в вышележащие центры, в симметричные структуры противоположной стороны, к передним рогам спинного мозга. Задние рога содержат афферентные нейроны, которые реагируют на болевые, температурные, тактильные, вибрационные, проприоцептивные раздражения.
Передние рога содержат нейроны (мотонейроны), дающие аксоны к мышцам, они являются эфферентными. Все нисходящие пути ЦНС двигательных реакций заканчиваются в передних рогах.
В боковых рогах шейных и двух поясничных сегментов располагаются нейроны симпатического отдела вегетативной нервной системы, во втором-четвертом сегментах – парасимпатического.
В составе СМ имеется множество вставочных нейронов, которые обеспечивают связь с сегментами и с вышележащими отделами ЦНС, на их долю приходится 97 % от общего числа нейронов спинного мозга. В их состав входят ассоциативные нейроны – нейроны собственного аппарата СМ, они устанавливают связи внутри и между сегментами.
Белое вещество СМ образовано миелиновыми волокнами (короткими и длинными) и выполняет проводниковую роль.
Короткие волокна связывают нейроны одного или разных сегментов спинного мозга.
Длинные волокна (проекционные) образуют проводящие пути спинного мозга. Они формируют восходящие пути, идущие к головному мозгу, и нисходящие пути, идущие от головного мозга.
Спинной мозг выполняет рефлекторную и проводниковую функции.
Рефлекторная функция позволяет реализовать все двигательные рефлексы тела, рефлексы внутренних органов, терморегуляции и т. д. Рефлекторные реакции зависят от места, силы раздражителя, площади рефлексогенной зоны, скорости проведения импульса по волокнам, от влияния головного мозга.
Рефлексы делятся на:
1) экстероцептивные (возникают при раздражении агентами внешней среды сенсорных раздражителей);
2) интероцептивные (возникают при раздражении прессо-, механо-, хемо-, терморецепторов): висцеро-висцеральные – рефлексы с одного внутреннего органа на другой, висцеро-мышечные – рефлексы с внутренних органов на скелетную мускулатуру;
3) проприоцептивные (собственные) рефлексы с самой мышцы и связанных с ней образований. Они имеют моносинаптическую рефлекторную дугу. Проприоцептивные рефлексы регулируют двигательную активность за счет сухожильных и позотонических рефлексов. Сухожильные рефлексы (коленный, ахиллов, с трехглавой мышцы плеча и т. д.) возникают при растяжении мышц и вызывают расслабление или сокращение мышцы, возникают при каждом мышечном движении;
4) позотонические рефлексы (возникают при возбуждении вестибулярных рецепторов при изменении скорости движения и положения головы по отношению к туловищу, что приводит к перераспределению тонуса мышц (повышению тонуса разгибателей и уменьшению сгибателей) и обеспечивает равновесие тела).
Исследование проприоцептивных рефлексов производится для определения возбудимости и степени поражения ЦНС.
Проводниковая функция обеспечивает связь нейронов СМ друг с другом или с вышележащими отделами ЦНС.
Спинальное животное – животное, у которого пересечен СМ, часто на уровне шеи, но функция большей части СМ сохраняется;
Сразу после перерезки СМ большинство его функций ниже места пересечения у спинального животного резко угнетаются. Через несколько часов (у крыс и кошек) или несколько дней, недель (у обезьян) большинство свойственных спинному мозгу функций восстанавливаются почти до нормы, обеспечивая возможность экспериментального исследования препарата.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом