
Фосфолипиды представляют собой сходные по структуре молекулы, состоящие из глицеринового скелета с фосфодиэфирными группами в положении С3, соединенные со спиртовыми полярными группами и двумя эстерифицированными глицерином жирными кислотами в положении С1’ и C2’. Природные фосфолипиды в положении C1’ содержат насыщенную жирную кислоту, а в положении C2’ — ненасыщенную жирную кислоту. С химической точки зрения самой простой формой фосфолипидов является фосфатидиловая кислота (ФК), в которой молекула спирта отсутствует. Эта молекула рассматривается как «фосфатидиловый» компонент более сложных фосфолипидов, название которых зависит от типа спиртовой группы. В клетках человека спиртовые группы состоят из азотистых оснований (холин, этаноламин, серин), глицерина или инозитола. В соответствии с этим фосфолипиды имеют названия: фосфатидилхолин (ФХ), фосфатидилэтаноламин (ФЭ), фосфатидилглицерин (ФГ), фосфатидилинозитол (ФИ) и фосфатидилглицерин (кардиолипин). Кардиолипин (КЛ) является уникальным фосфолипидом, состоящим из двух диэфирных фосфатных групп, соединенных молекулой глицерина. Химическая структура полярной «головы» определяет суммарный электрический заряд и ионное состояние фосфолипида. ФХ и ФЭ, имея отрицательно заряженную фосфатную группу и положительно заряженную аминогруппу, электрически нейтральны и называются нейтральными фосфолипидами, ФС, ФГ, ФК, ФИ и КЛ являются отрицательно заряженными или анионными фосфолипидами [McNeil H. P. и соавт., 1991].
Физические свойства фосфолипидов исследованы на модели мембранных систем, наиболее простой из которых является диспергирование липидов в жидкости. В этой ситуации фосфолипиды принимают одну из трех возможных конфигураций: мицеллярную, двухслойную или ламелларную (типичное состояние фосфолипидов биомембран) и гексагональную фазу. Способность фосфолипидов находиться в той или иной фазе зависит как от внутренних свойств молекул (природа «головной» группы, длина и степень насыщения жирной кислоты), так и внешних факторов (гидратация, температура, pH и ионная сила раствора). Присутствие дивалентных катионов, других липидов и белков также влияет на поведение фосфолипидов в системе за счет изменения формы их молекулы, которая в наиболее простых случаях зависит от соотношения площадей, занимаемых гидрофобными жирными кислотами и гидрофильными полярными «головными» группами.
Например, поскольку ФЭ имеет меньшую, чем ФХ «головную» группу и водородный мостик между фосфатной и аминогруппой, постольку уменьшается площадь молекулы в «головной» группе, и он спонтанно занимает гексагональную фазу. Другие фосфолипиды обычно занимают ламелларную фазу, но при изменении липидного микроокружения могут переходить в гексагональную. Ионы кальция индуцируют эту конфигурацию у КЛ за счет связывания с анионной группой, что уменьшает электростатическое отталкивание и площадь «головной» группы.
В клеточной мембране фосфолипиды образуют двойной слой, в котором гидрофобные цепи жирных кислот направлены внутрь мембраны, а гидрофильные полярные группы кнаружи. Мембранные белки прикрепляются периферически за счет полярных или ионных взаимодействий, или включаются в состав липидного слоя. Биологические мембраны обладают «жидкостными» свойствами, так как в пределах одного слоя отдельные липидные молекулы способны меняться местами с соседними со скоростью более миллиона раз в секунду. Обмен липидных молекул между слоями (flip-flop) является значительно более редким событием. Клеточная мембрана обладает выраженной асимметрией в плане распределения различных классов фосфолипидов во внешнем и внутреннем слоях. Холин-содержащие нейтральные фосфолипиды, такие как сфингомиелин и ФХ локализуются на внешней стороне мембраны в сочетании с небольшим количеством ФЭ. Внутренняя (цитозольная) часть ее состоит из небольшого количества ФХ и сфингомиелина, большого количества ФЭ, а также ФС и ФИ. Таким образом, в норме анионные фосфолипиды не присутствуют на внешней поверхности биомембран.
Читайте также: Запасательной функцией характеризуется ткань мышечная поперечнополосатая
У млекопитающих КЛ локализуется главным образом в сердечной ткани на внутренней мембране митохондрий, где он составляет не менее 20% от всех фосфолипидов. КЛ не выявляется на плазменных мембранах, на которых около 50—60% от общего пула фосфолипидов составляют сфингомиелин и ФХ, 20—30% — ФЭ, 10—15% ФС и менее 5% — ФИ. Сфингомиелин и ФХ адаптируют преимущественно ламелларную конфигурацию и, следовательно, вносят основной вклад в стабильность клеточной мембраны. Хотя ФЭ принимает преимущественно гексагональную фазу, он стабилизируется в клеточной мембране в присутствии других липидов. Однако некоторые специализированные функции клеточных мембран, например, экзоцитоз, требуют транзиторного перехода в другую, гексагональную фазу, и в этом процессе ФЭ, вероятно, играет важную роль.
Мембранная асимметрия фосфолипидов — универсальный феномен, характерный практически для всех клеток [Schroit A. J. и Zwaal R. F. A., 1991]. В настоящее время установлено, что поддержание асимметрии является активным процессом, реализующимся за счет активности АТФ и сульфгидрил-зависимого липидного насоса (аминофосфолипидтранслоказы), который перемещает аминофосфолипиды по направлению к внутренней мембране [Schroit A. J. и Zwaal R. F. A., 1991; Devaux P. F., 1992]. Активация приводит к увеличению движения фосфолипидов между слоями, что сопровождается потерей асимметрии. В частности, тромбоциты при стимуляции утрачивают нормальную асимметрию мембраны, что приводит к увеличению экспансирования анионного ФС. Этот процесс играет важную физиологическую роль в развитии локальной реакции свертывания крови. Кроме того, поверхностная экспозиция ФС (по крайней мере, на эритроцитах) является сигналом для быстрого выведения этих клеток из кровяного русла.
Отрицательно заряженные фосфолипиды создают поверхность, на которой происходит сборка ферментных комплексов двух основных реакций коагуляционного каскада. В одном из них (теназный комплекс) фактор X активируется комплексом фактора IXa и VIIIa, а в другом — протромбиназная реакция, происходит конверсия протромбина в тромбин ферментным комплексом, состоящим из фактора Xa и Va (протромбиназный комплекс). Взаимодействие факторов IXa, Xa и протромбина с липидной поверхностью происходит посредством образования кальций-зависимого мостика между остатками гамма-карбоксиглютаминовой кислоты этих белков и отрицательно заряженными полярными группами фосфолипидов. Связывание с липидной поверхностью приводит к увеличению локальной концентрации и эффективного расположения коагуляционных факторов, что способствует максимальной скорости протекания реакции. Любые вещества, мешающие сборке этих комплексов на фосфолипидной поверхности, в том числе антитела к фосфолипидам потенциально способны повышать уровень образования тромбина и нарушать свертывание крови.
Читайте также: Сколько надо ткани для обивки стула
Синтез фосфолипидов и триацилглицеролов тесно связаны
Начальные реакции синтеза триацилглицеролов и фосфолипидов совпадают и происходят при наличии глицерола и жирных кислот .
В реакциях биосинтеза можно выделить следующие события:
1. Образование глицерол-3-фосфата через диоксиацетонфосфат из глюкозы или при фосфорилировании свободного глицерола.
2.. Биосинтез фосфатидной кислоты – требует наличия глицерол-3-фосфата и жирных кислот. При связывании глицерол-3-фосфата с жирными кислотами синтезируется фосфатидная кислота.
Далее фосфатидная кислота может превращаться двумя путями – в ЦДФ-ДАГ или дефосфорилироваться до 1,2-ДАГ.
3. Синтез триацилглицерола (липогенез) – идет из 1,2-ДАГ после дефосфорилирования фосфатидной кислоты. Образованный 1,2-ДАГ ацилируется до ТАГ.
4. Синтез фосфолипидов . Сейчас рассматриваются два пути синтеза фосфолипидов.
- По одному пути 1,2-ДАГ не превращается в ТАГ, а связывается с этаноламином с образованием фосфатидилэтаноламина, либо с холином – образуется фосфатидилхолин.
- По другому пути, ЦДФ-ДАГ связывается либо с инозитолом, либо с серином с образованием соответственно фосфатидилинозитола или фосфатидилсерина. При декарбоксилировании фосфатидилсерина далее образуется фосфатидилэтаноламин, который может превратиться, в свою очередь, в фосфатидилхолин.
Синтезированный любым способом фосфатидилэтаноламин также способен взаимодействовать с серином и обратно образовывать фосфатидилсерин.
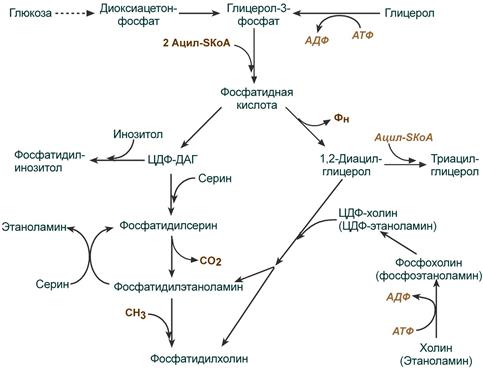
Общая схема реакций синтеза триацилглицеролов и фосфолипидов
ЦДФ-ДАГ, являясь активной формой фосфатидной кислоты, способен превращаться не только в фосфатидилинозитол, фосфатидилсерин, но и в другие фосфолипиды, например в кардиолипин .
Синтез фосфолипидов
В отличие от триацилглицеролов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды, как было сказано ранее, играют важную роль в структуре и функции клеточных мембран.
Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазматической сетке клетки.
Центральную роль в биосинтезе фосфолипидов играют L-диацилглицеролы (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфингомиелинов).
В этих биосинтезах принимает участие цитидинтрифосфат (ЦТФ), выполняющий роль активатора.
Основными компонентами мембран являются фосфатидилэтаноламин и фосфатидилхолин.
В процессе синтеза фосфатидилэтаноламина вначале происходит активация этаноамина под действием этаноламинкиназы:
Этаноламинфосфат реагирует затем с цитидинтрифосфатом (ЦТФ), в результате чего образуется цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (РРі). Эту реакцию катализирует фосфоэтаноламинцитидинтрансфераза:
Активированный ЦДФ–этаноламин при участии фермента этаноламинфосфаттрансферазы вступает в реакцию с L–диацилглицеролом с образованием фосфатидиламина:
L-диацилглицерол образуется при гидролизе фосфатидной кислоты. Аналогичные реакции ведут к образованию ЦДФ-холина.; последний вступает в реакцию с L-диацилглицеролом, в результате чего образуется фосфатидилхолин.
В синтезе фосфатидилэтаноламина и фосфатидилхолина важную роль играет ЦТФ. Подобно тому синтезу олиго- и полисахаридов, мы и здесь видим, каким образом нуклеотиды могут выполнять функцию переносчиков опреденных химических групп в обмене веществ клеток.
Читайте также: Укажите какие presets есть для быстрой настройки физических параметров ткани
Помимо синтеза фосфолипидов таким способом, в клетке они легко подвергаются взаимопревращениям. Фосфатидилэтаноламин является предшественником фосфатидилхолина. Так, в печени в результате последовательного переноса трех метильных групп от трех молекул S-аденозилметионина образуется фосфатидилхолин:
В переносе метильных групп важную роль играют тетрогидрофолевая кислота (витамин Вс) и метилкобаломин ( витамин В12).
Синтез фосфатидилсерина происходит другим путем с предварительным вовлечением фосфатидной кислоты:
Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина:
У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем:
Таким же путем образуется фосфатидилинозитол.
В биосинтезе сфингомиелина принимает участие церамид (N-ацетилсфингозин), который образуется при взаимодействии спирта сфингозина и ацетил-КоА. Сфингомиелин синтезируется в результате взаимодействия церамида с ЦДФ-холином:
Все выше синтезированные фосфолипиды поступают с помощью липидпереносящих белков из цитоплазмы к мембранам и участвуют в их построении.
Для синтеза фосфолипидов нужны липотропные факторы
Синтез фосфолипидов
Биосинтез фосфолипидов по сравнению с синтезом ТАГ имеет существенные особенности. Они заключаются в дополнительной активаци и компонентов ФЛ – фосфатидной кислоты или холина и этаноламина.
1 путь – «спасательный»
Благодаря этому пути холин и этаноламин используются повторно и не катаболизируют. Активация холина (или этаноламина ) происходит через промежуточное образование фосфорилированных производных с последующим присоединением ЦМФ. В следующей реакции фосфохолин (или фосфоэтаноламин) переносится на ДАГ. Этот путь особенно характерен для легких и кишечника, но идет и в других тканях.
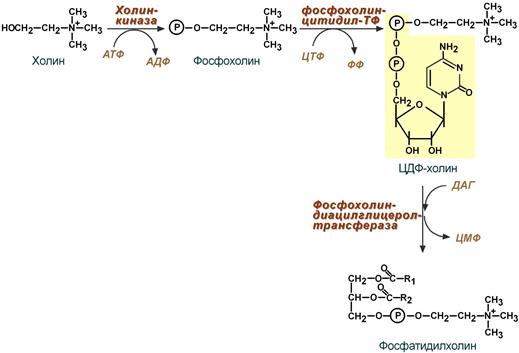
Реакции синтеза фосфолипидов
с использованием 1,2-ДАГ на примере фосфатидилхолина
2 путь – основной, синтез de novo
Здесь холин (или этаноламин) не встраиваются в готовом виде, а образуются уже в молекуле фосфолипида.
Активация фосфатидной кислоты заключается в присоединении к ней ЦМФ с образованием ЦДФ-ДАГ. Далее к нему присоединяется шестиатомный спирт инозитол или серин с образованием фосфатидилинозитола и фосфатидилсерина . Синтезированный фосфатидилсерин подвергается декарбоксилированию с образованием фосфатидилэтаноламина . Последний метилируется при участии S-аденозилметионина в фосфатидилхолин .
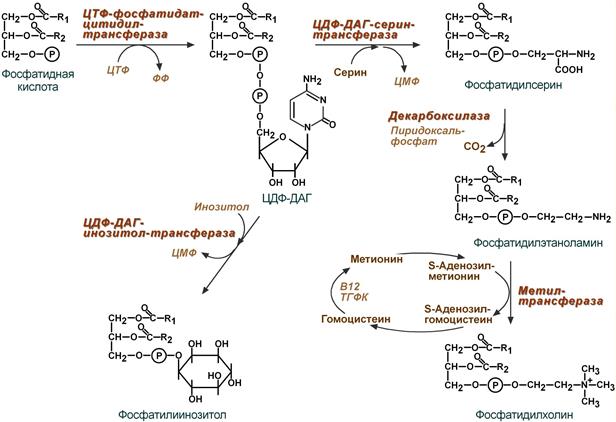
Реакции синтеза фосфолипидов
с использованием фосфатидной кислоты
3 путь – обратное превращение
Между фосфатидилэтаноламином и серином может происходить реакция с образованием в результате реакции фосфатидилсерина и свободного этаноламина .
Липотропные вещества
Все вещества, способствующие синтезу ФЛ и препятствующие синтезу ТАГ, и способные предотращать жировую инфильтрацию печени, называются липотропными факторами . К ним относятся:
1. Структурные компоненты фосфолипидов: полиненасыщенные жирные кислоты, инозитол, серин, холин, этаноламин.
2. Метионин – в виде S-аденозилметионина является донором метильных групп для синтеза холина и фосфатидилхолина.
- пиридоксин (В6), способствующий образованию ФЭА из ФС.
- цианкобаламин (В12) и фолиевая кислота, участвующие в реакциях обмена серина, глицина и метионина при ресинтезе метионина из гомоцистеина. Следовательно они, хотя и не напрямую, но необходимы для синтеза фосфатидилхолина.
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом
- Правообладателям
- Политика конфиденциальности
Мастерица © 2023
Информация, опубликованная на сайте, носит исключительно ознакомительный характер