
С тех пор процессы пролиферации и дифференцировки клеток ушли далеко вперед, создав настоящее чудо — вас, человека. У растения жизнь начинается точно так же — с одной маленькой клетки, из которой в дальнейшем будут развиваться ткани и органы самых разных форм. Главная заслуга роста растения принадлежит образовательной ткани.
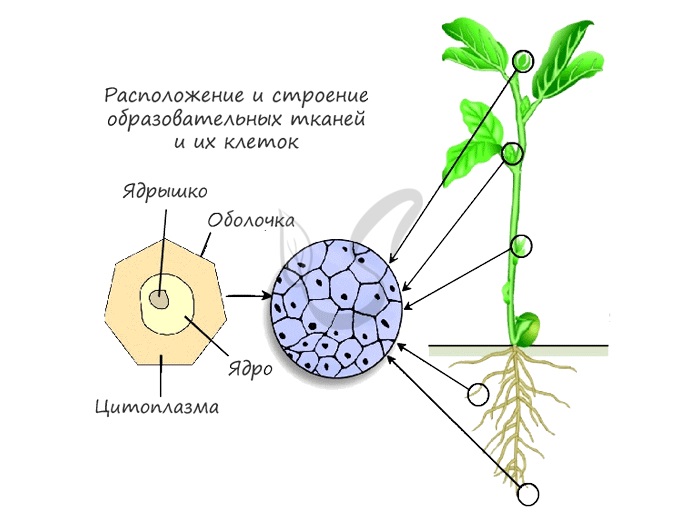
Как вы видите, на картинке схематично изображено месторасположение образовательной ткани. Главным образом это:
- Кончик побега — конус нарастания в почках
- Кончик корня — зона деления, прикрытая корневым чехликом для защиты
- Камбий — обеспечивает рост растения в ширину
- Основание междоузлий и черешков листьев — это также зоны активного роста растения
Именно в этих местах и происходит деление клеток и рост растения. Важно отметить, что сезонные изменения активности клеток камбия являются причиной возникновения годичных колец древесины. Внешний вид годичных колец обусловлен хронологической закономерностью: весной больше образуется проводящей ткани (более тонкая и рыхлая внутри), а осенью — механическая (толстая, более твердая). Именно поэтому годичные кольца на спиле дерева выглядят как чередование колец, отличающихся друг от друга.
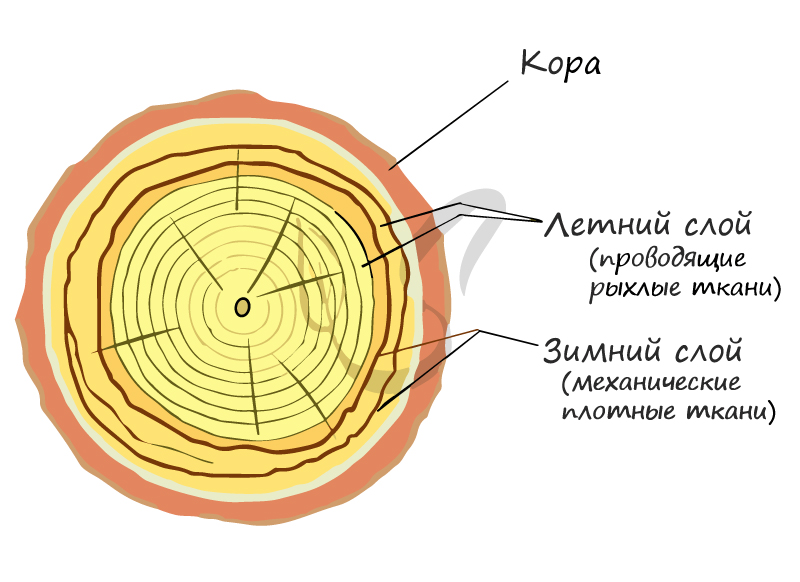
На внешний вид годичных колец оказывают весьма сильное влияние условия внешней среды. Так, при дефиците трофического компонента (питательных веществ), к примеру, у растений, растущих на болоте, годичные кольца выглядят тоньше своих обычных размеров.
Ветер также оказывает существенное влияние: при его постоянном действии происходит перераспределение древесины по стволу. Оказывая действие на крону, ветер смещает центр тяжести дерева, что сказывается на его нижележащих отделах. Они начинают компенсаторно утолщаться для предотвращения слома дерева. При постоянно дующем ветре ствол сильно искривляется, а форма кроны становится флагообразной.

Тема камбия и форм стволов растений весьма занимательна, и все-таки мы должны разобраться в строении самой образовательной ткани. Она представлена живыми мелкими быстро делящимися клетками с относительно крупным ядром. Объем цитоплазмы небольшой, она вязкая по консистенции, оболочка клетки тонкая. Это уязвимые клетки, которые растение оберегает по-своему, подобно тому, как животные оберегают только что появившееся потомство.

Другое название образовательных тканей — меристемы (с др.-греч. — «μεριστός» — делимый). По времени возникновения различают первичные и вторичные меристемы.
Первичные меристемы — закладываются в эмбриогенезе
1) Вставочные меристемы (интеркалярные) — в виде отдельных участков в зоне активного роста в разных частях растения. Такие ткани можно найти в основании междоузлий у злаков, черешков листьев у многих растений. У злаковых наблюдается быстрый рост стебля за счет множественного расположения данной ткани на стебле — «вставочный рост».
2) Прокамбий — основа будущего камбия, перицикла, окружающего проводящие ткани в один или несколько слоёв (у голосеменных). В корнях перицикл является корнеродным слоем, так как в корне с него начинается формирование осевого цилиндра, наружным слоем которого он является. В нём закладываются придаточные и боковые корни, что имеет принципиальное значение для формирования корневой системы растения.
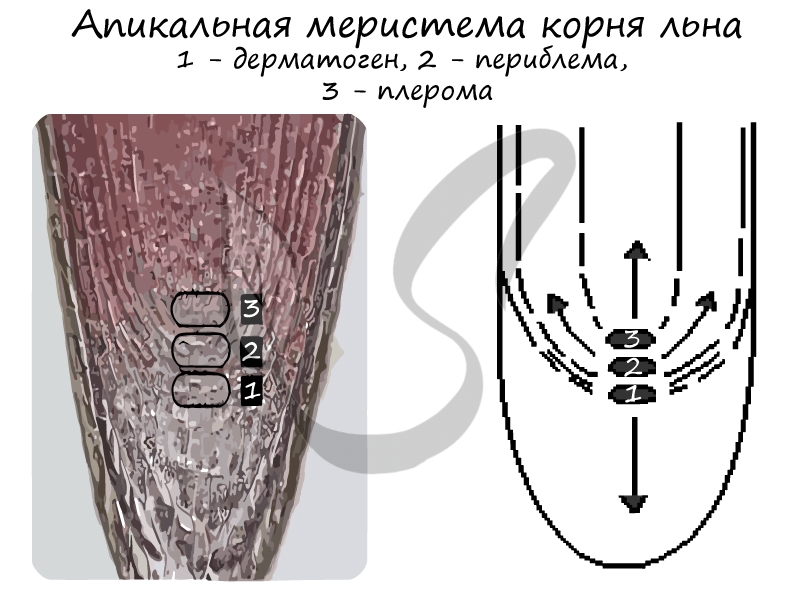
3) Верхушечные (апикальные) — формируются на верхушках стеблей и кончиках корней. В периферической части корня различают три слоя:
- Дерматоген — в дальнейшем преобразующийся в первичную покровно-всасывающую ризодерму (эпиблему или ризодерму)
- Периблема — образующая ткани первичной коры
- Плерома — внутренний слой ткани центрального осевого цилиндра
Вторичные меристемы — закладываются в постэмбриональном развитии
Камбий и феллоген (пробковый камбий) — занимают боковое положение по отношению к оси органа, обеспечивают рост вширь. Растения часто повреждаются, их задевают животные, нарушая целостность тканей и органов. На этот случай в группе вторичных меристем есть раневые меристемы, дающие начало защитной ткани в местах повреждения растения.

Топографическая классификация меристем
Спешу заверить, это отнюдь не сложная классификация, которой нужно бояться. Речь пойдет о взгляде на те же образовательные ткани с другой стороны. В переводе с греч. τόπος — место. Мы рассмотрим меристемы в соответствии с их месторасположением на растении.
- Верхушечная или апикальная (лат. apex — вершина) — расположена на кончике корня и конусе нарастания побега
- Боковая или латеральная (лат. latus — бок): камбий – обеспечивает рост стебля и корня в толщину
- Краевая или маргинальная (лат. margo — край) меристема даёт начало листовой пластинке
- Вставочная или интеркалярная (лат. inter — между и calaris — вставочный, добавочный) — расположена преимущественно у основания стеблевых междоузлий между зонами дифференцированных тканей.
© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: Ткань костюмка описание тянется или нет
МОРФОЛОГИЯ РАСТЕНИЙ — Т. А. Сауткина — 2012
ГЛАВА 3. ТКАНИ РАСТЕНИЙ
3.1. Понятие о тканях. Принципы классификации тканей
В процессе эволюции, которая была связана с приспособлением к условиям суши прикрепленных талломных организмов, дифференцировка тела растений сопровождалась усложнением и их внутренней структуры. Возникли группы клеток, которые выполняют определенную функцию. У древнейших, известных науке наземных растений — риниофитов — уже присутствовали комплексы клеток разных типов. Еще в 1671 г. Н. Грю предложил называть группы схожих клеток тканями.
Н. Грю, а также М. Мальпиги определили основные морфологические особенности разных групп клеток. В 1807 г. Г. Линк (1707—1851) ввел понятие о паренхимных и прозенхимных клетках, а те клетки, которые разрастаются более или менее равномерно по всем направлениям и диаметр которых примерно одинаков или соотношение длины и ширины не превышает двух, получили название паренхимных. Такие клетки имеют изодиаметрическую или таблитчатую форму. Клетки, у которых длина более чем в два раза превышает ширину, были названы прозенхимными. Обычно они входят в состав проводящей и механической ткани. Это значит, что с морфологической точки зрения растения состоят всего из двух типов клеток. Но в дифференцированном теле высших растений возникли группы клеток, которые похожи не только по форме, но и по функциям. Такие устойчивые комплексы клеток, схожие по происхождению, строению, выполняемым функциям и занимающие определенное место в теле растений, называются тканями. Существуют разные типы тканей и различные подходы к их классификации. При характеристике тканей можно использовать как один какой-нибудь их важный признак, так и весь комплекс признаков, подчеркивающих не только строение клеток, формирующих ткань, но и ее функциональные особенности.
С учетом одного ведущего признака ткани подразделяют:
1) по форме клеток, которые образуют ткань, — ткани паренхимные (основная паренхима, эпидермис) и прозенхимные (проводящие элементы, механические волокна).
2) физиологическому состоянию клеток — ткани мертвые и живые. В мертвых тканях отмирает цитоплазма, но клеточная оболочка сохраняется и продолжает играть важную роль (трахеиды, сосуды, древесинные волокна). В клетках живых тканей есть цитоплазма и наблюдается циклоз.
3) степени дифференциации клеток — ткани недифференцированные (меристематические) с высокой тотипотентностью — способностью к реализации всей генетической программы организма и ткани постоянные, дифференцированные по функциям (покровные, основные, проводящие и др.).
4) времени и особенностям образования — первичные, вторичные и третичные. Первичные ткани образуются из первичных, зародышевых или апикальных меристем (эпидермис, колленхима). Вторичные — из вторичных меристем или из постоянных тканей (вторичные проводящие ткани, камбий, феллоген). Третичные ткани образуются у высоковозрастных растений как комплекс вторичных покровных и расположенных между ними различных отмерших тканей (ритидом). Вторичные ткани обеспечивают вторичное утолщение и характерны в основном для голосеменных и покрытосеменных двудольных растений. Они отсутствуют у мхов, хвощей, плаунов, подавляющего большинства папоротников, а также у покрытосеменных однодольных растений.
5) по степени сложности выделяют ткани простые, состоящие из однотипных клеток (колленхима, паренхима, ризодерма), и сложные, включающие клетки нескольких типов (ксилема, флоэма, перидерма).
В определенной степени к классификации имеют отношение все перечисленные подходы. Однако наиболее рациональной и признанной является классификация тканей на основе анатомо-физиологических принципов, т. е. на основании строения тканей и выполняемых ими функций. Впервые такую классификацию тканей предложил в конце XIX в. Ф. Габерланд (1828—1878). Он выделил 9 систем с учетом максимальной физиологической активности тканей. В настоящее время некоторые авторы придерживаются этой классификации, но в большинстве случаев морфологи используют упрощенную классификацию, согласно которой все ткани высших растений объединяются в 6 типов:
1) образовательные, или меристемы;
2) покровные, или пограничные;
4) механические, или арматурные (опорные);
6) выделительные структуры (сборная группа).
Наряду с главной функцией нередко ткани могут выполнять и дополнительные, что позволяет рассматривать их как полифункциональные.
3.2. Анатомо-морфологические и функциональные особенности меристематических тканей
Тело растений формируется в результате деятельности образовательных тканей, или меристем (от греч. meristos — делимый). Именно за счет меристем растения растут, образуют новые корни, побеги, цветки.
Цитологические признаки меристематических тканей связаны с функциями, которые они выполняют. Клетки апикальных меристем мелкие, более или менее изодиаметричные, многогранные, тесно сомкнутые, без межклетников. Они имеют тонкую первичную оболочку, которая содержит небольшое количество целлюлозы и способна растягиваться. Клетки меристемы характеризуются высокой метаболической активностью. В них находится густая цитоплазма с довольно крупным (относительно небольших размеров клетки) ядром. В клетках содержится много рибосом, но незначительное количество пластид; митохондрии слабо дифференцированы. Эргастические вещества (вещества запаса), как правило, отсутствуют. Вакуоли мелкие, малозаметные.
Клетки меристем способны к активному митотическому делению в течение всей жизни растения или достаточно длительное время, чем отличаются от образовательных тканей животных. Меристемы сохраняются столь длительный срок благодаря тому, что имеют одну (хвощи) или несколько (покрытосеменные) так называемых инициальных клеток. Эти клетки имеют вид многогранников (обычно граней 14). Инициальные клетки меристем сохраняют способность к делению в течение всей жизни растения и производят только меристематические клетки. Они делятся неограниченное количество раз в разных плоскостях. Как правило, каждое последующее деление происходит параллельно одной из граней 14-гранника. Остальные, дочерние клетки, образуют основную меристему, клетки которой делятся ограниченное число раз и, в конце концов, превращаются в постоянные ткани — проводящие, покровные, механические, основные и т. д.
Читайте также: Выкройки попугаев из ткани
В зависимости от происхождения и времени появления в онтогенезе различают две группы меристем: первичные и вторичные.
Первичные меристемы берут начало с момента деления зиготы и начальных стадий образования зародыша (предзародыша). До тех пор, пока предзародыш не станет довольно крупным шаровидным телом, все клетки его обладают меристематической активностью и делятся в разных направлениях. С момента образования боковых элементов зародыша — семядолей — происходит первая локализация меристемы на апикальном (от лат. apex — вершина) полюсе стебля. Между семядолями дифференцируется конус нарастания стебля (апекс), представленный первичной апикальной (верхушечной) меристемой. На противоположной стороне дифференцируется конус нарастания корня, также образованный первичной меристемой. Именно за счет деятельности апикальных меристем корень и побег нарастают в длину. При ветвлении каждый боковой корень и боковой побег имеют свои апикальные меристемы и инициали, что позволяет этим органам расти в длину. Апикальные меристемы обусловливают так называемый первичный рост осевых органов не только в длину, но и в толщину.
Апикальные меристемы корня и стебля представляют собой довольно сложно дифференцированные ткани (рис. 13). В 1868 г. немецкий ботаник И. Ганштейн (1822—1880) при изучении апекса корня обнаружил в нем три достаточно четко выделяющиеся зоны. Он назвал их дерматоген, периблема и плерома. Так возникла теория гистогенов — тканей-образователей (от греч. histos — ткань и genos — происхождение). Согласно представлениям, И. Ганштейна каждый из гистогенов имеет свою инициальную клетку. И. Ганштейн полагал, что из каждого слоя верхушечной меристемы корня образуются различные элементы корня (топографические зоны). Так как дерматоген однослойный, а его клетки способны делиться только антиклинально (перпендикулярно поверхности органа), из него образуется первичная покровная ткань корня — ризодерма.
Рис. 13. 1. Строение верхушечной (апикальной) меристемы конусов нарастания (А—Б). II. Схема дифференцировки апексов согласно теориям, И. Ганштейна (В),
Дж. Будера и А. Шмидта (Г), А. Фостера (Д): А—корень подсолнечника обыкновенного (Helianthus annuus); Б—стебель водяной сосенки (Hippuris vulgaris): 1—корневой чехлик, 2—инициальные клетки апикальной меристемы, 3—дерматоген, 4—периблема, 5—плерома, 6—калиптроген; 7—протодерма, 8—туника, 8а—протодерма, 9—корпус, 10—мантия, 11—зона центральных материнских клеток, 12—фланговая зона, 13—медуллярная зона

Периблема состоит из нескольких слоев меристематических клеток, которые делятся в различных направлениях и образуют особую топографическую зону корня — первичную кору. Она состоит из паренхимной ткани. Самый мощный слой меристемы апекса представлен плеромой. Из него формируется важнейшая топографическая зона корня — центральный цилиндр, или стель. Центральный цилиндр имеет сложное строение. На ранних этапах развития в нем из периферических клеток плеромы образуются особые типы меристем — трицикл и прокамбий, а также развивается паренхима центрального цилиндра. Таким образом, перицикл и прокамбий — это производные апикальной меристемы.
Теория гистогенов после открытия получила широкое распространение и на первых порах сыграла большую роль в изучении формирования тканей у различных растений в ходе онтогенеза. Однако, как показали дальнейшие исследования, она не была универсальной. Не у всех растений обнаруживалось в апексе корня три гистологические зоны. После критических замечаний А. де Бари и других авторов интерес к теории гистогенов ослабел.
В 1924—1928 гг. немецкие ботаники Дж. Будер и А. Шмидт разработали теорию туники и корпуса (рис. 13, Б). Они установили, что в апексе стебля покрытосеменных растений меристема дифференцирована только на две зоны. Туника выражена слабо, самый наружный ее слой дает эпидермис, остальные слои образуют первичную кору. Центральный цилиндр формируется из корпуса. Согласно теории туники и корпуса, роль конуса нарастания побега велика не только в период формирования осевого органа — стебля, но и при образовании его элементов — листьев и почек.
Теория гистогенов и теория туники и корпуса не противоречат друг другу, они показывают только специфичность меристем осевых органов, которые находятся в разных условиях существования. Если размеры и форма конуса нарастания корня на разных этапах онтогенеза постоянные, то эти параметры у конуса нарастания стебля изменяются и имеют возрастные и сезонные различия.
В 1938 г. американский ботаник А. Фостер при изучении конуса нарастания стебля гинкго двулопастного (отдел Голосеменные) указал, что конус нарастания делится на мантию и корпус, которые образуются апикальными меристемами. В то же время он обнаружил, что корпус неоднороден по величине клеток, активности их деления, строению оболочек, развитию вакуолей, окрашиванию гистологическими красителями. Это связано с тем, что у голосеменных есть группа инициалей, расположенных не только на верхушке конуса нарастания (апикальные клетки), но и в глубже лежащих слоях.
Читайте также: Убираем следы от утюга с ткани
Самый верхний слой апикальных инициальных клеток образует мантию. Она состоит из более или менее однородных меристематических клеток. Именно этот слой соответствует тунике (по теории Дж. Будера и А. Шмидта), но у голо- и покрытосеменных клетки этого слоя делятся по-разному: у голосеменных — периклинально (параллельно поверхности органа) и антиклинально (перпендикулярно поверхности органа), а у покрытосеменных, деления только антиклинальные.
Под апикальными меристематическими клетками расположена зона центральных материнских клеток корпуса, они образуют медуллярную и фланговую зоны. Фланговая зона участвует в образовании прокамбия, части первичной коры и листовых бугорков. Клетки медуллярной зоны формируют сердцевину стебля.
Таким образом, теория гистоцитологической зональности А. Фостера дополняет теорию туники и корпуса Дж. Будера и А. Шмидта и пытается объяснить особенности образования элементов побега различным строением и неодинаковой меристематической активностью клеток в его апексе. Все указанные теории дают представление о дифференцировке апексов (конуса нарастания, точки роста) корня и стебля, что, несомненно, важно для понимания процессов формирования первичного анатомического строения осевых органов, но ни одна из этих теорий не является универсальной.
Прокамбий и перицикл как производные апикальных меристем являются первичными меристемами и имеют ряд специфических особенностей.
Клетки прокамбия прозенхимные, в поперечном сечении многоугольные с хорошо развитыми крупными вакуолями. Прокамбий не имеет инициальных клеток, образует продольные тяжи и является васкулярной меристемой (от лат. vasculum — уменьшительное от vas — сосуд), из него формируются проводящие ткани.
Перицикл — особая образовательная ткань, которая хорошо выражена в корне. Как и прокамбий, он не имеет инициальных клеток. Чаще всего перицикл состоит из одного ряда клеток таблитчатой формы, вытянутых параллельно поверхности корня.
К первичным меристемам относятся также интеркалярные, или вставочные, меристемы. Они отделяются от верхушечной меристемы, между ними развиваются зоны постоянных, дифференцированных тканей. В интеркалярных меристемах отсутствуют инициальные клетки, поэтому они функционируют ограниченное время, вскоре прекращают делиться и превращаются в постоянные ткани. Интеркалярные меристемы находятся в основании междоузлий побега (особенно долго они сохраняются у злаков), тычиночных нитей, листовых зачатков, черешков листьев, цветоносов. Они характерны для тех растений, у которых часто повреждаются апексы или на верхушке закладываются зачатки цветков либо соцветий.
К вторичным меристемам относятся камбий, феллоген, раневые меристемы. Вторичный рост растений, или вторичное утолщение, связано с появлением и деятельностью вторичной меристематической ткани — камбия. Камбий может образоваться из прокамбия, перицикла или паренхимы, способной восстановить меристематическую функцию. Клетки камбия таблитчатой формы, вытянуты параллельно поверхности осевых органов. Камбий — васкулярная меристема, ее клетки делятся периклинально (параллельно поверхности) и дают начало вторичным проводящим тканям. Для камбия характерно наличие инициалей. Инициальные клетки несколько различаются по форме и дают начало разным элементам вторичных проводящих тканей: из паренхиматозных клеток инициалей образуются элементы лучевой паренхимы, а из прозенхимных (веретеновидных) — проводящие элементы.
Феллоген, или пробковый камбий, — особая вторичная меристема, образующаяся из клеток эпидермы, субэпидермальных клеток паренхимы первичной коры, а также из перицикла. Феллоген дает начало перидерме — вторичной покровной ткани.
По положению в теле растения прокамбий, камбий, а также перицикл являются латеральными (боковыми) меристемами.
К вторичным относятся также раневые меристемы, которые возникают при повреждении тканей и органов растения. В результате паренхимные клетки вокруг раны начинают активно делиться в разных направлениях, образуется наплыв, который называется каллусом (от лат. callus — толстая кожа, толстая мозоль). Иногда из паренхимных клеток формируется феллоген, который затягивает рану пробкой.
Клетки различных меристем могут делиться в разных направлениях. Если межклеточная перегородка закладывается под прямым углом к поверхности органа, такое деление называется антиклинальным. За счет подобных делений образуются однослойные покровные ткани. Деления, при которых вновь образующиеся перегородки между клетками параллельны ближайшей поверхности органа, называются периклинальными. Они особенно характерны для латеральных меристем; за счет таких делений обычно происходит утолщение осевых органов. В том случае если перегородка закладывается касательно окружности, деление называется тангентальным (тангенциальным).
В процессе дифференцировки клеток меристемы в постоянные ткани можно наблюдать два типа роста: симпластический и интрузивный (интерпозиционный). При симпластическом типе рост оболочек соседних клеток происходит согласованно, их плазмодесменные связи не нарушаются. При интрузивном типе роста подобная согласованность отсутствует, и одни клетки внедряются в пространство, образовавшееся между другими клетками.
Биологическая библиотека — материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
© 2018-2022 Все права на дизайн сайта принадлежат С.Є.А.