
Мышечные ткани — это ткани, для которых способность к сокращению является главным свойством. Мышечные ткани составляют активную часть опорно-двигательного аппарата (пассивной частью являются кости, соединения костей).
Общими свойствами всех мышечных тканей является сократимость и возбудимость. К данной группе тканей относятся гладкая, поперечнополосатая скелетная и поперечнополосатая сердечная мышечные ткани. Клетки мышечной ткани имеют хорошо развитый цитоскелет, содержат много митохондрий.

Гладкая (висцеральная) мускулатура
Эта мышечная ткань встречается в стенках внутренних органах (бронхи, кишечник, желудок, мочевой пузырь), в стенках сосудов, протоках желез. Эволюционно является наиболее древним видом мускулатуры.
Состоит из веретенообразных миоцитов — коротких одноядерных клеток. Между клетками имеются межклеточные контакты — нексусы (лат. nexus — связь). Благодаря нексусам возбуждение, возникшее в одной клетке, волнообразно распространяется на все остальные клетки.

Гладкая мышечная ткань отличается своей способностью к длительному тоническому напряжению, что очень важно для работы внутренних органов (к примеру мочевого пузыря), сокращается медленно, практически не утомляется. Скелетная мышечная ткань, которую мы изучим чуть позже, такой способностью не обладает — сокращается и утомляется быстро.
Осуществляется сокращение с помощью клеточных органоидов — миофиламентов, которые расположены в клетке хаотично и не имеют такой упорядоченной структуры, как миофибриллы в скелетной мускулатуре (все познается в сравнении, уже скоро мы их тоже изучим).
Особо заметим, что в гладкой мышечной ткани миофиламенты собираются в миофибриллы только во время сокращения. У таких временных миофибрилл не может быть регулярной организации, а значит ни у таких миофибрилл, ни у гладких миоцитов не может быть поперечной исчерченности.
Гладкая мышечная ткань сокращается непроизвольно (неподвластна воле человека). Работа гладких мышц обеспечивается вегетативной (автономной) нервной системой. К примеру невозможно по желанию сузить или расширить бронхи, кровеносные сосуды, зрачок.

Гладкая мышечная ткань называется неисчерченной, так как не обладает поперечной исчерченностью, характерной для поперечнополосатых скелетной и сердечной мышечных тканей.
Скелетная (поперечнополосатая) мышечная ткань
Скелетная мышечная ткань образует диафрагму (дыхательную мышцу), мускулатуру туловища, конечностей, головы, голосовых связок.
В отличие от гладкой мускулатуры, скелетная образована не отдельными одноядерными клетками, а длинными многоядерными волокнами, имеющими до 100 и более ядер — миосимпластами. Миосимпласт (греч. sim — вместе + plast — образованный) представляет совокупность слившихся клеток, имеет длину от нескольких миллиметров до нескольких сантиметров (соответствует длине мышцы).
Внутри миосимпласта находится саркоплазма, снаружи миосимпласт покрыт сарколеммой. Сократительные элементы — миофибриллы (лат. fibra — волоконце) — длинные тяжеобразные органеллы в миосимпласте (около 1400).

Характерная черта данной ткани — поперечная исчерченность, выражающаяся в равномерном чередовании светлых и темных полос на мышечном волокне. Это происходит потому, что границы саркомеров в соседних миофибриллах совпадают, вследствие чего все волокно приобретает поперечную исчерченность. Теперь самое время изучить микроскопическую основу мышцы — саркомер.
Саркомер (от греч. sarco — мясо (мышца) + mere — маленький)
Саркомер — элементарная сократительная единица поперечнополосатых мышц, структурная единица миофибриллы. В состав саркомера (и миофибриллы в целом) входят миофиламенты (лат. filamentum — нить) двух типов, которые обеспечивают сократимость мышечной ткани.
Саркомер состоит из актиновых (тонких) и миозиновых (толстых) филаментов, которые образованы главным образом белками актином и миозином. Сокращение происходит за счет взаимного перемещения миофиламентов: они тянутся навстречу друг другу, саркомер укорачивается (и мышца в целом).

Источником энергии для сокращения служат молекулы АТФ. К тому же невозможно представить сокращение мышц без участия ионов кальция: именно они связываются с тропонином, что приводит к изменению конформации тропомиозина (тропонин и тропомиозин — регуляторные белки между нитями актина), за счет чего становится возможно соединение актина и миозина. При сокращении мышц выделяется тепло (сократительный термогенез).
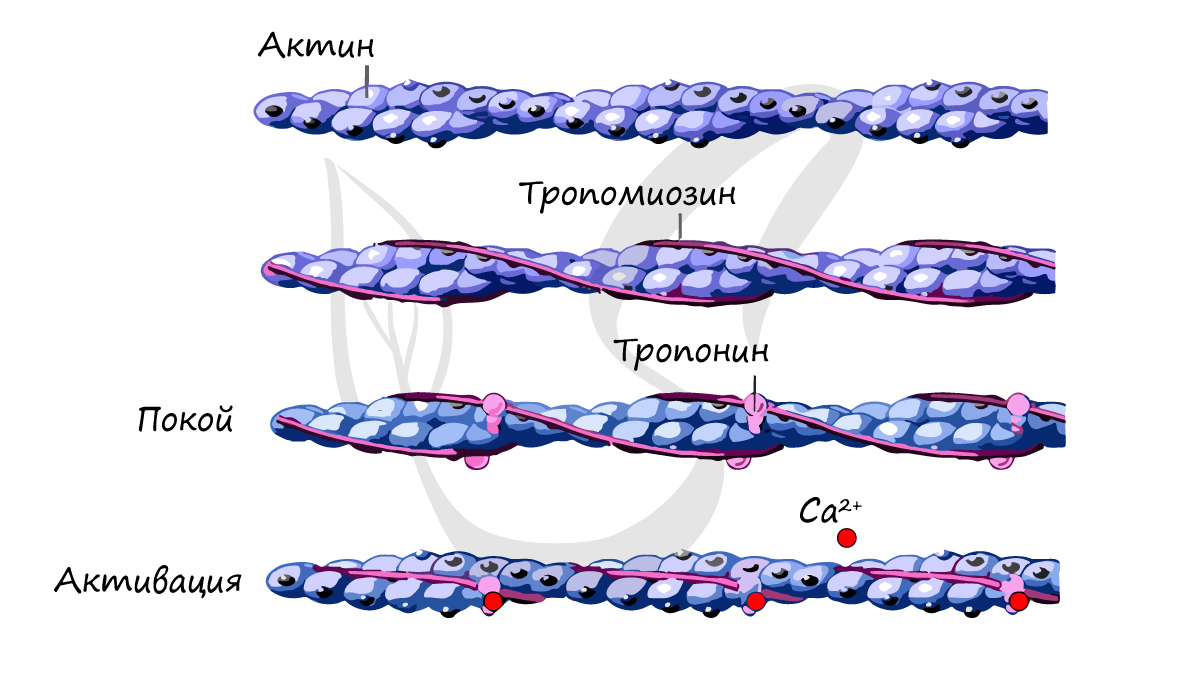
Замечу, что трупное окоченение (лат. rigor mortis) — посмертное затвердевание мышц — связано именно с ионами кальция, которые устремляются в область низкой концентрации (в саркоплазму миосимпласта), способствуя связыванию актина и миозина.
Читайте также: Ткань синтетическое волокно состав
После смерти в мышце перестает синтезироваться АТФ, ее уровень быстро снижается. Как следствие этого перестает функционировать Ca-АТФаза — насос, выкачивающий ионы Ca из саркоплазмы в саркоплазматический ретикулум (мембранная органелла мышечных клеток (сходная с ЭПС), в которой запасаются ионы Ca).
В саркоплазме повышается концентрация ионов Ca — замыкаются мостики между актином и миозином, однако разомкнуться они уже не могут, в связи с чем наблюдается стойкая мышечная контрактура (лат. contractura — стягивание, сужение): конечности очень сложно разогнуть или согнуть.

Вернемся к скелетным мышцам. Имеется еще ряд важных моментов, о которых нужно знать.
В процесс возбуждения вовлекается изолированно один миосимпласт, соседние миосимпласты (волокна) не возбуждают друг друга, в отличие от гладких миоцитов, где возбуждение предается между соседними клетками через нексусы. Скелетные мышцы сокращаются быстро и быстро утомляются (у гладких мышц фазы сокращения и расслабления растянуты во времени, мало утомляются) .
Скелетные мышцы сокращаются произвольно: они подконтрольны нашему сознанию. К примеру, по желанию мы можем изменить скорость движения руки, темп бега, силу прыжка. Мышцы покрыты фасцией, крепятся к костям сухожилиями, и, сокращаясь, приводят в движение суставы.

Сердечная поперечнополосатая мышечная ткань
Сердечная мышечная ткань образует мышечную оболочку сердца — миокард (от др.-греч. μῦς «мышца» + καρδία — «сердце»). Миокард — средний слой сердца, составляющий основную часть его массы. При работе сердечная мышечная ткань не утомляется.

Сердечная мышечная ткань состоит из кардиомиоцитов — одиночных клеток, имеющих поперечную исчерченность. Соединяясь друг с другом, кардиомиоциты образуют функциональные волокна.
Этот тип мышечной ткани удивительным образом сочетает свойства двух предыдущих, изученных нами, тканей (возбудимость, сократимость) и имеет одно новое уникальное свойство — автоматизм.
Автоматизм — способность сердечной мышечной ткани возбуждаться и сокращаться самопроизвольно, без влияний извне. Это легко можно подтвердить, наблюдая сокращения изолированного сердца лягушки в физиологическом растворе: сокращения сердца в нем будут продолжаться несколько десятков минут после отделения сердца от организма.

Места контактов соседних кардиомиоцитов — вставочные диски (в их составе находятся нексусы), благодаря которым возбуждение одной клетки передается на соседние, таким образом волнообразно охватываются возбуждением и сокращаются новые участки миокарда.
Большое число контактов между кардиомиоцитами обеспечивает высокую эффективность и надежность проведения возбуждения по миокарду. Сокращается эта ткань непроизвольно, не утомляется.
На рисунке или микропрепарате узнать данную ткань можно по центральному положению ядер в клетках, поперечной исчерченности, наличию вставочных дисков и анастомозов (греч. anastomosis — отверстие) — мест соединений боковых поверхностей функциональных волокон (кардиомиоцитов).

В норме возбуждение проводится по проводящей системе сердца от предсердий к желудочкам (однонаправленно). Участок сердечной мышцы, в котором генерируются импульсы, определяющие частоту сердечных сокращений — водитель сердечного ритма.
Автоматизм возможен благодаря наличию в миокарде особых пейсмекерных (англ. pacemaker — задающий ритм) клеток, которые также называют водителями ритма. Они спонтанно генерируют нервные импульсы, которые охватывают весь миокард, в результате чего осуществляется сокращение. Именно благодаря водителям ритма сердце лягушки продолжает биться, будучи полностью отделенным от тела.
Ответ мышц на физическую нагрузку
Физические нагрузки приводят к гипертрофии мышц (от др.-греч. ὑπερ- чрез, слишком + τροφή — еда, пища) — в них увеличивается количество мышечных волокон, объем мышечной массы нарастает.

В условиях гиподинамии (от греч. ὑπό — под и δύνᾰμις — сила), то есть пониженной активности, мышцы уменьшаются вплоть до полной атрофии (греч. а – «не» + trophe – питание). В худшем случае волокна мышечной ткани перерождаются в соединительную ткань, после чего пациент становится обездвиженным.

Необходимо отметить, что сердечная мышечная ткань также дает ответную реакцию на чрезмерную нагрузку: сердце увеличивается в размере, нарастает масса миокарда. Причиной могут быть генетические заболевания, повышенное артериальное давление. Гипертрофия сердца — состояние, требующее вмешательства врача и наблюдения за пациентом.
Читайте также: Физиологическая регенерация эпителиальных тканей
В большинстве случае гипертрофия сердца обратима, а у спортсменов наблюдается так называемая физиологическая гипертрофия (вариант нормы).

Происхождение мышц
Мышцы развиваются из среднего зародышевого листка — мезодермы.

© Беллевич Юрий Сергеевич 2018-2022
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Из каких источников не развиваются мышечные ткани
Развитие мышц.
Мышцы тела должны рассматриваться с точки зрения их развития и функции, а также топографии систем и групп, в которые они складываются.
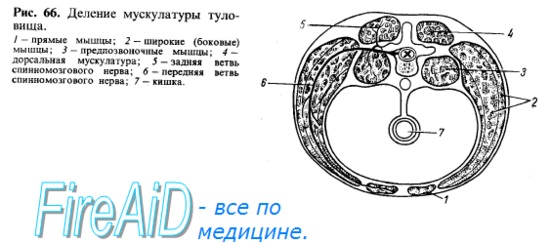
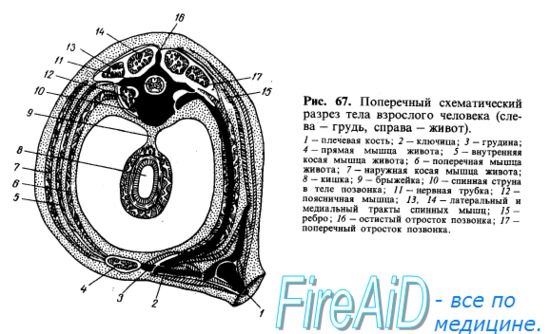
Мышцы туловища развиваются из залегающей по бокам хорды и мозговой трубки дорсальной части мезодермы, которая разделяется на первичные сегменты, или сомиты. После выделения скелеротома, идущего на образование позвоночного столба, оставшаяся дорсомедиальная часть сомита образует миотом, клетки которого (миобласты) вытягиваются в продольном направлении, сливаются друг с другом и превращаются в дальнейшем в симпласты мышечных волокон. Часть миобластов дифференцируется в особые клетки — миосателлиты, лежащие рядом с симпластами. Миотомы разрастаются в вентральном направлении и разделяются на дорсальную и вентральную части. Из дорсальной части миотомов возникает спинная (дорсальная) мускулатура туловища, а из вентральной — мускулатура, расположенная на передней и боковой сторонах туловища и называемая вентральной (рис. 66, 67).
В каждый миотом (миомер) врастают ветви соименного спинномозгового нерва (невромера). Соответственно делению миотома на 2 части от нерва отходят 2 ветви, из которых дорсальная (задняя) входит в дорсальную часть миотома, а вентральная (передняя) — в вентральную. Все происходящие из одного и того же миотома мышцы снабжаются одним и тем же спинномозговым нервом. Соседние миотомы могут срастаться между собой, но каждый из сросшихся миотомов удерживает относящийся к нему нерв. Поэтому мышцы, происходящие из нескольких миотомов (например, прямая мышца живота), иннервируются несколькими нервами. Первоначально миотомы на каждой стороне отделяются друг от друга поперечными соединительнотканными перегородками, myosepta (рис. 68). Такое сегментированное расположение мускулатуры туловища у низших животных остается на всю жизнь. У высших же позвоноч.ых и у человека благодаря более значительной дифференцировке мышечных масс сегментация значительно сглаживается, хотя следы ее и остаются как в дорсальной (короткие мышцы перекидывающиеся между позвонками), так и в вентральной мускулатуре (межреберные мышцы и прямая мышца живота). Часть мышц, развившихся на туловище, остается на месте, образуя местную, аутохтонную мускулатуру (autos — тот же самый, chthon, греч. — земля). Другая часть в процессе развития перемещается с туловища на конечности. Такие мышцы называются трункофугальными (truncus — ствол, туловище, fugo — обращаю в бегство). Наконец, третья часть мышц, возникнув на конечностях, перемещается на туловище. Это трункопетальпые мышцы (рею — стремлюсь). На основании иннервации всегда можно отличить аутохтонную (т. е. развивающуюся в данном месте) мускула гуру от сместившихся в эту область других мышц-пришельцев
Мускулатура конечностей образуется из мезенхимы почек конечностей и получает свои нервы из передних ветвей спинномозговых нервов при посредстве плечевого и пояснично-крестцового сплетений. У низших рыб (селахий) из миотомов туловища вырастают мышечные почки, которые разделяются на два слоя, расположенные с дорсальной и вентральной сторон скелета плавника. Подобным же образом у наземных позвоночных мышцы по отношению к зачатку скелета конечности первоначально располагаются дорсально и вентрально (разгибатели и сгибатели). При дальнейшей дифференцировке зачатки мышц передней конечности разрастаются и в проксимальном направлении (трункопетальные мышцы) и покрывают аутохтонную мускулатуру туловища со стороны груди и спины (mm. pectorales major et minor, m. latissimus dorsi). Кроме этой первичной мускулатуры передней конечности, к поясу верхней конечности присоединяются еще трункофугальные мышцы, т. е. производные вентральной мускулатуры. служащие для передвижения и фиксации пояса и переместившиеся на него с головы (mm. trapezius и sternocleidomastoideus) и с туловища (mm. rhomboideus, levator scapulae, serratus anterior, subclavius, omohyoideus). У пояса задней конечности вторичных мышц не развивается, так как он неподвижно связан с позвоночным столбом. Сложная дифференцировка мышц конечностей наземных позвоночных, в особенности у высших форм, объясняется функцией конечностей, превратившихся в сложные рычаги, выполняющие различного рода движения.
Мышцы головы возникают отчасти из головных сомитов, а главным образом из мезодермы жаберных дуг. Висцеральный аппарат у низших рыб состоиг из сплошного мышечного слоя (общий сжиматель), который делится по своей иннервации на отдельные участки, совпадающие с мета-мерным расположением жаберных дуг: 1-й жаберной (мандибулярной) дуге соответствует V пара черепных нервов (тройничный нерв), 2-й жаберной (гиоидной) дуге — VII пара (лицевой нерв), 3-й жаберной дуге — IX пара (языког лоточный нерв). Остальная часть общего сжимателя снабжается ветвями X пары (блуждающий нерв). Сзади общего сжимателя обособляется пучок, прикрепляющийся к поясу верхней конечности (трапециевидная мышца). Когда с переходом из воды на сушу у низших позвоночных прекратилось жаберное дыхание- приспособленное для жизни в воде, мышцы жаберного аппарата (висцеральные) распространились на череп, где превратились в жевательные и мимические мышцы, но сохранили свою связь с теми частями скелета, которые возникли из жаберных дуг. Поэтому жевательные мышцы, возникающие из челюстной дуги и мышцы дна рта, располагаются и прикрепляются на нижней челюсти и иннервируются тройничным нервом (V пара). Из мускулатуры, соответствующей 2-й жаберной дуге, происходит главным образом подкожная мускулатура шеи и головы, иннер-вируемая лицевым нервом (VII пара).
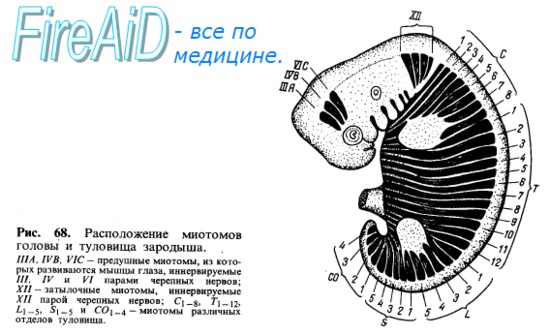
Мышцы, возникающие из материала обеих жаберных дуг, имеют двойное прикрепление и двойную иннервацию, например двубрюшная мышца, переднее брюшко которой прикрепляется к нижней челюсти (иннервация из тройничного нерва), а заднее — к подъязычной кости (иннервация из лицевого нерва). Висцеральная мускулатура, иннервируемая IX и X парами черепных нервов, у наземных позвоночных частью редуцируется, частью идет на образование мышц глотки и гортани. Трапециевидная мышца Теряет всякую связь с жаберными дугами и становится исключительно мышцей пояса верхней конечности. У млекопитающих от нее отщепляется в виде отдельной части грудино-ключично-сосцевидная мышца. Задняя ветвь блуждающего нерва, иннер-вирующая трапециевидную мышцу, превращается у высших позвоночных в самостоятельный черепной нерв — п. accessorius. Так как мозговой череп во всех своих частях представляет неподвижное образование, то на нем ожидать развития мышц нельзя. Поэтому на голове встречаются только некоторые остатки мускулатуры, образовавшейся из головных сомитов. К числу их нужно отнести мышцы глаза, происходящие из так называемых предушных миотомов (иннервация от III, IV и VI пар черепных нервов) (см. рис. 68).
Затылочные миотомы вместе с передними туловищными миотомами обычно образуют путем вентральных отростков особую поджаберную или подъязычную мускулатуру, лежащую под висцеральным скелетом. За счет этой мускулатуры, проникающей кпереди до нижней челюсти, происходят у наземных позвоночных мышцы языка, снабжаемые в силу своего происхождения из затылочных сомитов комплексом нервных волокон, образующих подъязычный нерв, который только у высших позвоночных стал настоящим черепным нервом. Остальная часть подъязычной мускулатуры (ниже подъязычной кости) представляет собой продолжение вентральной мускулатуры туловища, иннервируемой от передних ветвей спинномозговых нервов. Таким образом, для понимания расположения и фиксации мышц надо учитывать, кроме их функции, также и развитие (см. рис. 66, 67).
- Свежие записи
- Балкон в многоквартирном доме: является ли он общедомовым имуществом?
- Штраф за остекление балкона в 2022: что это и как избежать наказания
- Штраф за мусор с балкона: сколько заплатить за выбрасывание окурков
- Оформление балконного окна: выбираем шторы из органзы
- Как выбрать идеальные шторы для маленькой кухни с балконом